


-
二叠纪—三叠纪之交(2.52亿年前)生物大灭绝(Permian-Triassic Mass Extinction,PTME)是地球生命自显生宙以来遭受最残酷的浩劫(Sepkoski,1984;Erwin,1994;Chen and Benton,2012)。大灭绝后,后生生物大量消失殆尽。相反,蓝细菌、硫酸盐还原菌等多种微生物群落在浅海碳酸盐岩沉积区中迅速爆发(Luo et al.,2013;Heindel et al.,2018;Chen et al.,2019),形成广布的、具有地层对比意义的微生物岩/礁建造。同时,微生物岩生态系统也为生物在大灭绝后最早的复苏提供温床(Foster et al.,2018)。通过国内外学者的研究,使得人们对微生物岩的岩石组构特征、形成过程和机制以及微生物矿化作用等方面有了全面的了解(Kershaw et al.,2012;Chen et al.,2022;Wu et al.,2022,2024)。
但是,二叠纪末大灭绝后微生物岩的形成过程中仍旧存在一些亟待解决的悖论问题。比如,广泛分布的微生物岩建造代表了大灭绝后微生物的爆发,理论上有利于海洋生产力的提高,而地质记录却表明,大灭绝后的海洋古生产力偏低,微生物岩中总有机碳(Total Organic Carbon,TOC)含量极低(Algeo et al.,2013;Tang et al.,2017;Chen et al.,2022)。贵州省罗甸县田湾剖面记录了二叠纪—三叠纪之交的微生物岩建造,该剖面微生物岩富含多种微生物和后生生物化石,同时保存了大量不同大小的黑色泥晶球粒(peloids)或颗粒结构(Adachi et al.,2004)。然而,它们的分类、成因机制及其有机质来源尚不清楚。
Macintyre(1985)根据物质来源的不同,将球粒划分为三类,即粪便颗粒、碎屑颗粒和原位沉积颗粒,涵盖无机成因和有机成因。而Flügel(2004)提出了球粒的九种亚类,分别对应多样化的成因机制和形成环境。微生物岩中的球粒可能是蓝细菌等微生物钙化作用后的有机残留物,也可能是无机化学沉淀的产物(Alexandersson,1972;Samankassou et al.,2005;Broughton,2023)。微生物岩内部广泛分布蓝细菌和其他细菌,以及生物碎屑起源的多种球粒结构,球粒与致密的微晶方解石共同构建了微生物岩/礁(Zhou and Pratt,2019)。作为类似环境的产物,现代环境的冷泉钙华中也有大量球粒的报道,球粒由矿化的胞外聚合物(Extracellular Polymeric Substances,EPS)、无定形碳酸钙(Amorphous Calcium Carbonate,ACC)以及单水方解石(monohydrocalcite,MHC)微晶组成,微生物可以利用EPS为碳酸钙成核提供位点,控制ACC转变为MHC,形成球粒的核心结构(Broughton,2023)。此外,Samankassou et al.(2005)还发现白垩纪球粒源自分解磨损的微晶生物壳体,二者之间具有几乎相同的元素组成。总之,前人揭示地质历史时期球粒的起源往往与微生物群落有关。
因此,为探究二叠纪末大灭绝后微生物爆发与有机质埋藏的关系,本文深入解剖产自广西田湾剖面微生物岩的泥晶球粒结构,重建其沉积过程,探究其有机质来源,厘清由微生物产生的有机质和长时间滞留在海水中的有机质分别对泥晶微球粒形成过程的影响机制,旨在解决二叠纪末大灭绝后海洋中微生物爆发与古生产力低下的矛盾,以及正确理解大灭绝后微生物岩生态系统中碳循环机制。
-
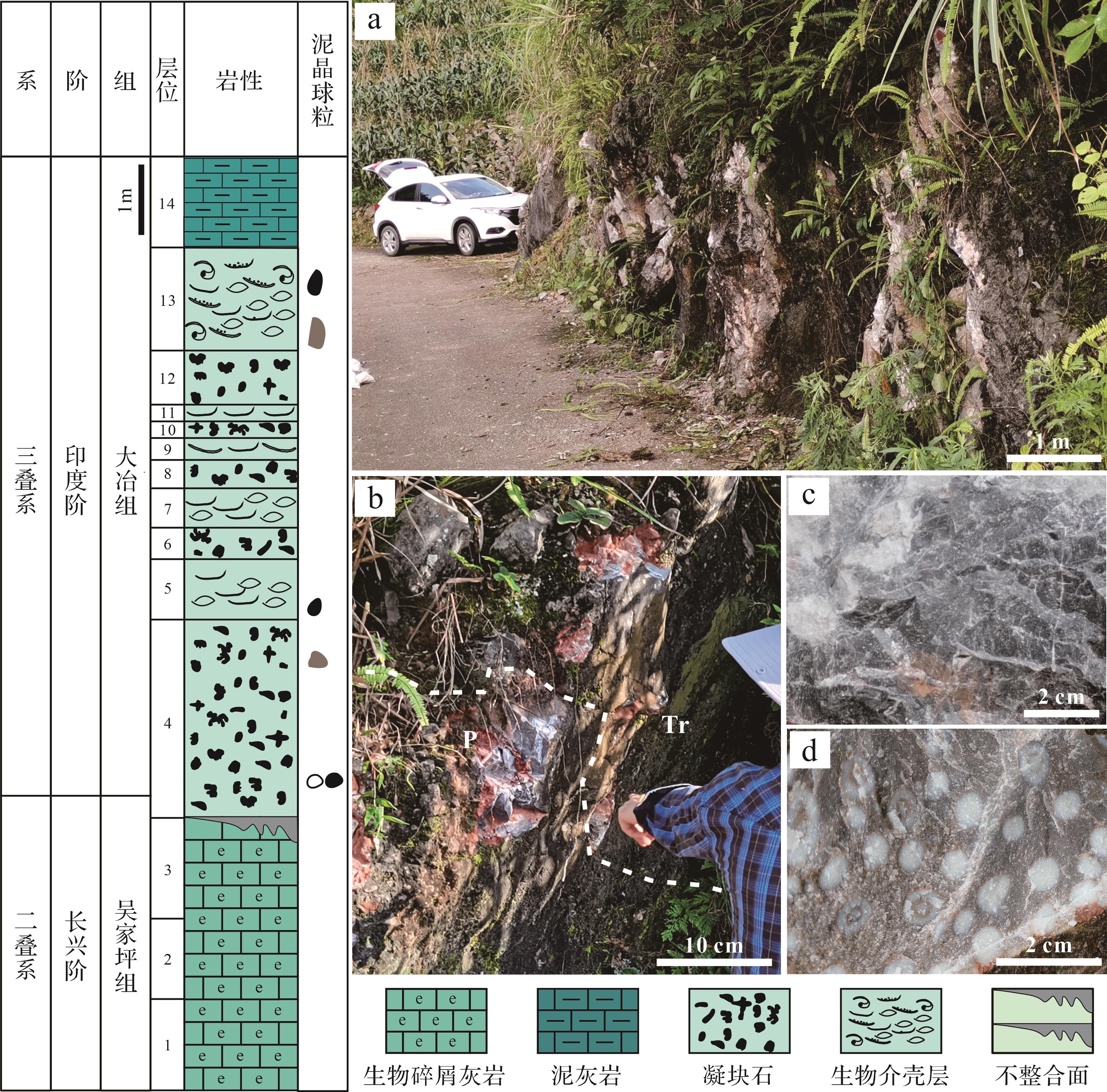
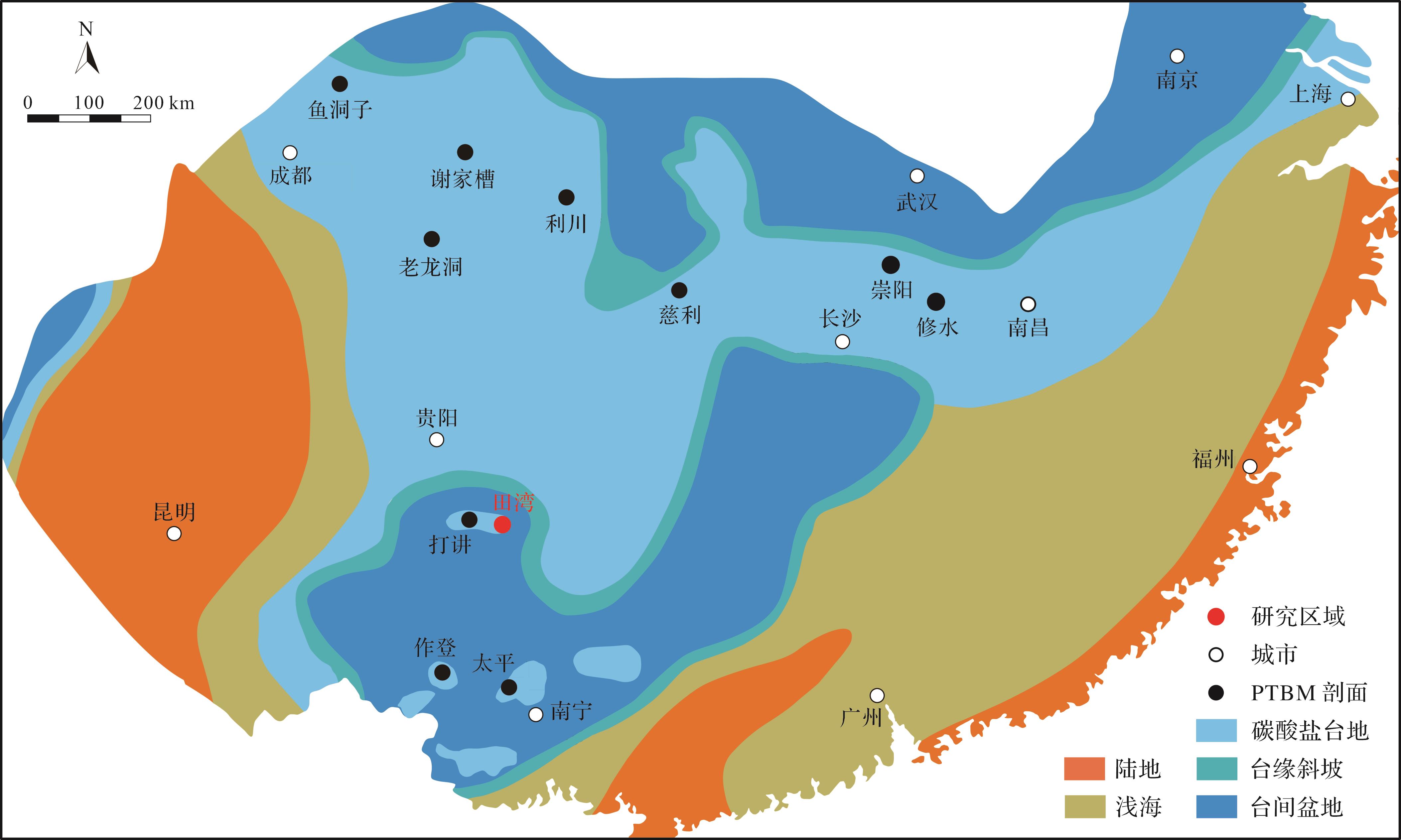
研究剖面位于中国西南部贵州省罗甸县以北约25 km处的田湾村(图1)。在二叠纪—三叠纪之交,该地区处于南盘江盆地腹地孤立的大贵州滩浅水台地边缘。受多期次构造运动叠加影响,南盘江盆地内部形成复杂的断裂系统,台地—盆地分异显著(冯增昭等,1997)。PTME后,微生物岩在全球低纬度地区的碳酸盐岩沉积体系中广泛发育,二叠纪—三叠纪界线(Permian-Triassic Boundary,PTB)通常置于该微生物岩建造的底部(Chen et al.,2022)。在华南地区南盘江盆地内部孤立碳酸盐台地之上,发育大量微生物岩。

图 1 华南早三叠纪古地理图,显示田湾剖面的古地理背景(底图据冯增昭等,1997修改)
Figure 1. Early Triassic paleogeographical map of South China showing the setting of the Tianwan section (base map modified from Feng et al., 1997)
田湾剖面发育的下三叠统归为大冶组,其底部发育厚约10.5 m的微生物岩,距微生物岩底部3.4 m和8.0 m处产出牙形石Hindeodus parvus及Isarcella estachei分子(Adachi et al.,2004),它们是三叠纪最早期的两个牙形石带的命名化石种(Yuan et al.,2018),尽管在田湾剖面上H. parvus的最低层位出现在微生物岩下部,但是对产自华南地区所有剖面的牙形石物种首现面的综合对比发现,H. parvus的最低层位在更多剖面中出现在微生物岩的底部(Wu et al.,2017,2022;Chen et al.,2022)。因此,将PTB置于微生物岩的底部。
微生物岩的下伏地层为上二叠统(乐平统)吴家坪组生物碎屑灰岩,它与微生物岩建造呈平行不整合接触(图2b)。微生物岩本身由凝块岩组成,偶夹生物介壳层透镜体。微生物岩的上覆地层为下三叠统下部薄层泥质灰岩(图2a)。

图 2 田湾剖面微生物岩序列的岩性柱状图以及典型岩性类型的野外露头
Figure 2. Lithostratigraphy of the Tianwan section showing microbialite succession, and field outcrop photographs
吴家坪组生物碎屑灰岩为颗粒支撑,颗粒间孔隙由亮晶方解石胶结,属于颗粒灰岩,内部包含丰富的后生生物化石,如珊瑚、介形虫、有孔虫、苔藓虫、棘皮动物、钙质藻类等,生物多样性较高,最顶部发育成层的珊瑚礁(图2d),灭绝事件发生后,后生生物大量灭绝,仅有少量物种在微生物岩中存活。
微生物岩的类型相对单一,主要由凝块石构成,其内部产丰富的化石,以蓝细菌等微生物化石为主,岩石组构以灰泥支撑为特色;其内部间隔分布多个颗粒支撑的生物介壳层透镜体,粒间孔隙多被灰泥或亮晶方解石充填,含有少量介形虫、有孔虫、微管虫、双壳、腹足等(图2c)化石,生物多样性明显降低(Adachi et al.,2004)。此外,不同类型的泥晶微球粒在中下部凝块石和顶部生物介壳层中广泛发育,它们具有多样的形态结构,特征差异显著,内部有机质含量普遍较高。
-
利用光学显微镜、电子显微镜及其能谱分析、拉曼光谱等分析手段对产自田湾剖面微生物岩中的泥晶微球粒进行细致的原位微区分析,探究球粒成因与生物过程的联系。首先,利用蔡司偏光显微镜对岩石薄片进行观察,统计不同类型球粒结构的粒径大小,并对球粒内部有机质的分布进行荧光成像分析,对荧光响应显著的区域进行共聚焦拉曼光谱测试和面扫,确定有机质的分布特征,并根据矿物特征峰的强度拟合生成矿物高峰光谱图像。同时,利用HITACHI-SU8010场发射扫描电子显微镜(Field Emission Scanning Electron Microscope,FE-SEM)对岩石样品新鲜断口进行观察,并利用能量色散X射线光谱(Energy-Dispersive X-ray Spectroscopy,EDS)进行元素成分半定量分析,重建生物化石和泥晶球粒结构的元素组成和分布特征。所有实验在中国地质大学(武汉)地质微生物与环境全国重点实验室完成。
随后,对球粒粒径和EDS元素成分百分数进行数理统计分析,计算其数值分布范围、置信区间、中值和均值,代表球粒粒径大小和元素组成分布情况。对产自微生物岩的泥晶球粒、生物化石和介壳碎片外部泥晶套的有机质进行拉曼频谱分析,有机质的拉曼光谱通常具有特定峰值系列:D1(1 340~1 350 cm⁻¹)、D2(1 600~1 620 cm⁻¹)、D3(1 510 cm⁻¹)、D4(1 245 cm⁻¹)和G(1 600 cm⁻¹),有机质在1 200~1 610 cm-1波段峰形组成差异明显(田野和田云涛,2020)。利用WITec Project Five 5.1 Plus软件对拉曼频谱数据进行背景去除和降噪处理,采用洛伦兹分峰拟合法拟合拉曼光谱,温度低于340 ℃的有机质,均可拟合出五个峰,分别是D1、D2、D3、D4、G;在340 ℃~600 ℃区域内,光谱最多可识别出D1、D2、D3、G峰等四个峰段,该温度范围内D4峰不明显;温度高于650 ℃后,有机质完全石墨化,仅有G峰信号保留(Kouketsu et al.,2014;田野和田云涛,2020)。拟合完成后利用表1公式计算有机质的成岩烘烤温度。参考其中碳质材料拉曼光谱的峰强、峰位、峰宽等信息恢复有机质成岩烘烤温度(Kouketsu et al.,2014),重建不同物质的成岩烘烤过程。
表 1 有机质拉曼温度计算公式
Table 1. Calformula for organic Raman temperature
公式编号 公式 适用温度范围/℃ 公式来源 描述 1 T=-2.15(FWHM.D1)+478 150~400 Kouketsu et al.,2014 基于D1峰的半峰宽(FWHM.D1)计算的温度 2 T=-6.78(FWHM.D2)+535 150~400 Kouketsu et al.,2014 基于D2峰的半峰宽(FWHM.D1)计算的温度 3 T=-445×R2+641 330~650 Beyssac et al.,2002 使用面积比参数R2[D1/(D1+D2+G)]计算温度 注: 公式1误差为±30 ℃,公式2与公式3误差为±50 ℃,单独使用某个公式受不同拉曼光谱参数对成岩烘烤温度的敏感性干扰,联合标定结果更加准确。 -
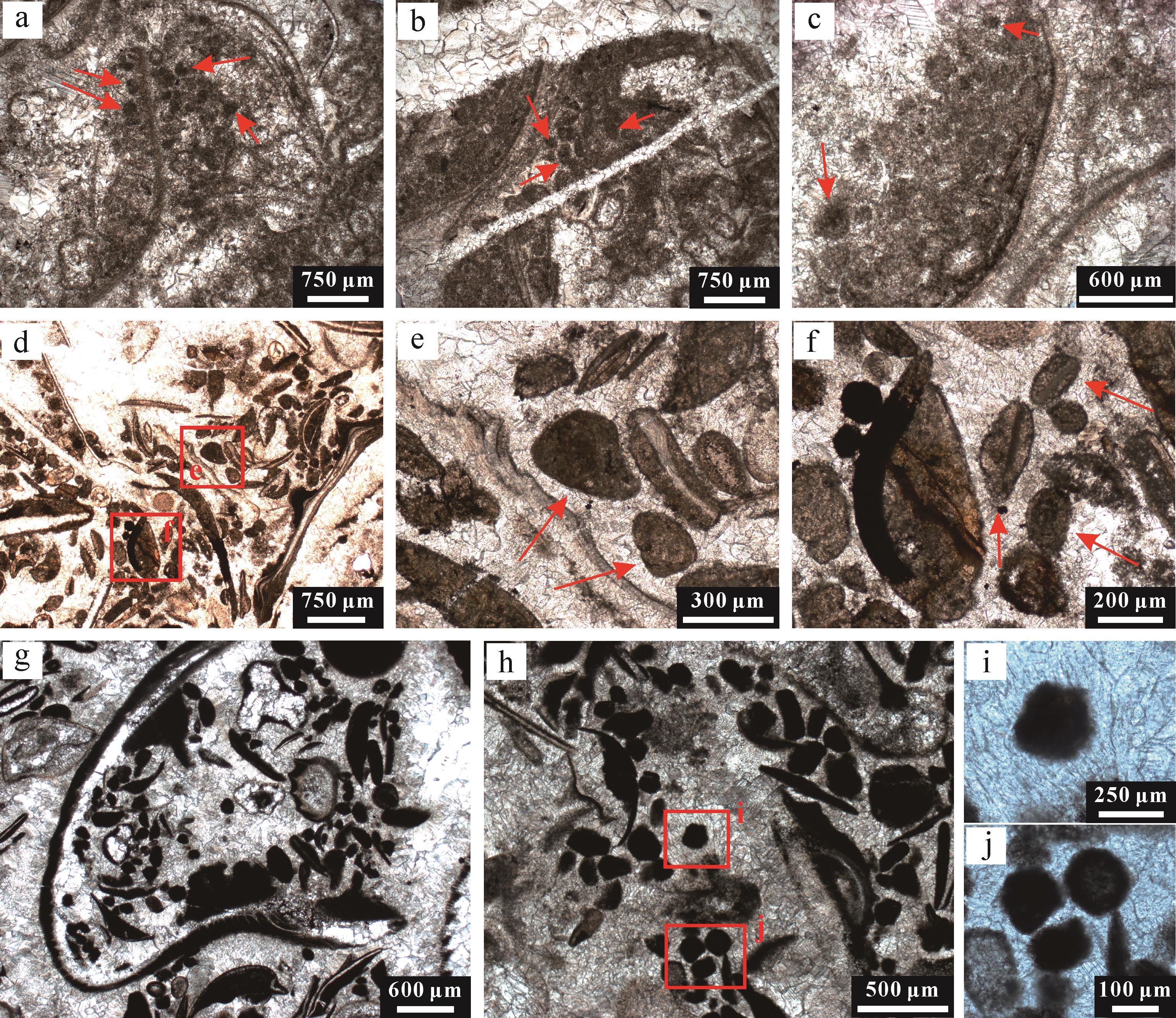
华南地区三叠系底部微生物岩的岩石组构类型包括叠层石、凝块石、树枝石和均一石。田湾微生物岩主要由凝块石构成,未见叠层石和树枝石构造。其底部发育多条橘黄色缝合线,含有大量以伊利石、蒙脱石为主的黏土矿物(图3c)。凝块石厚度从几十厘米到几米不等,发育不规则的亮晶结构块体,随机散布于灰泥基质中,组成凝块结构。微生物岩底部主要发育纹层状凝块石,上部则以“脑纹状”凝块石为主,其内部凝块结构明显。局部发育显著的穹窿状构造。

图 3 田湾剖面微生物岩中观及微观特征
Figure 3. Meso⁃microstructures of microbialites in the Tianwan section
中观—微观尺度上,凝块石由不规则的浅色亮晶团块与深色微晶凝块组成,亮晶团块由部分白云石化的粗晶方解石组成,而微晶团块则为细晶方解石。浅色与深色区域的接触边界不规则但清晰(图3a)。凝块结构为毫米至厘米级,主要由白色亮晶方解石团块组成,方解石空腔内充填大小不一的灰黑色圆形—椭圆形泥晶凝块结构(图3d,e),周围可见黄色泥晶填充物(图3c)。亮晶方解石团块内部,可见大量微米级囊泡结构集中分布,具有亮晶核部与泥晶边部,形态不规则且尺度较小(图3f)。不同尺度的凝块融合起来,组成凝块石的不规则网格格架(图3a)。
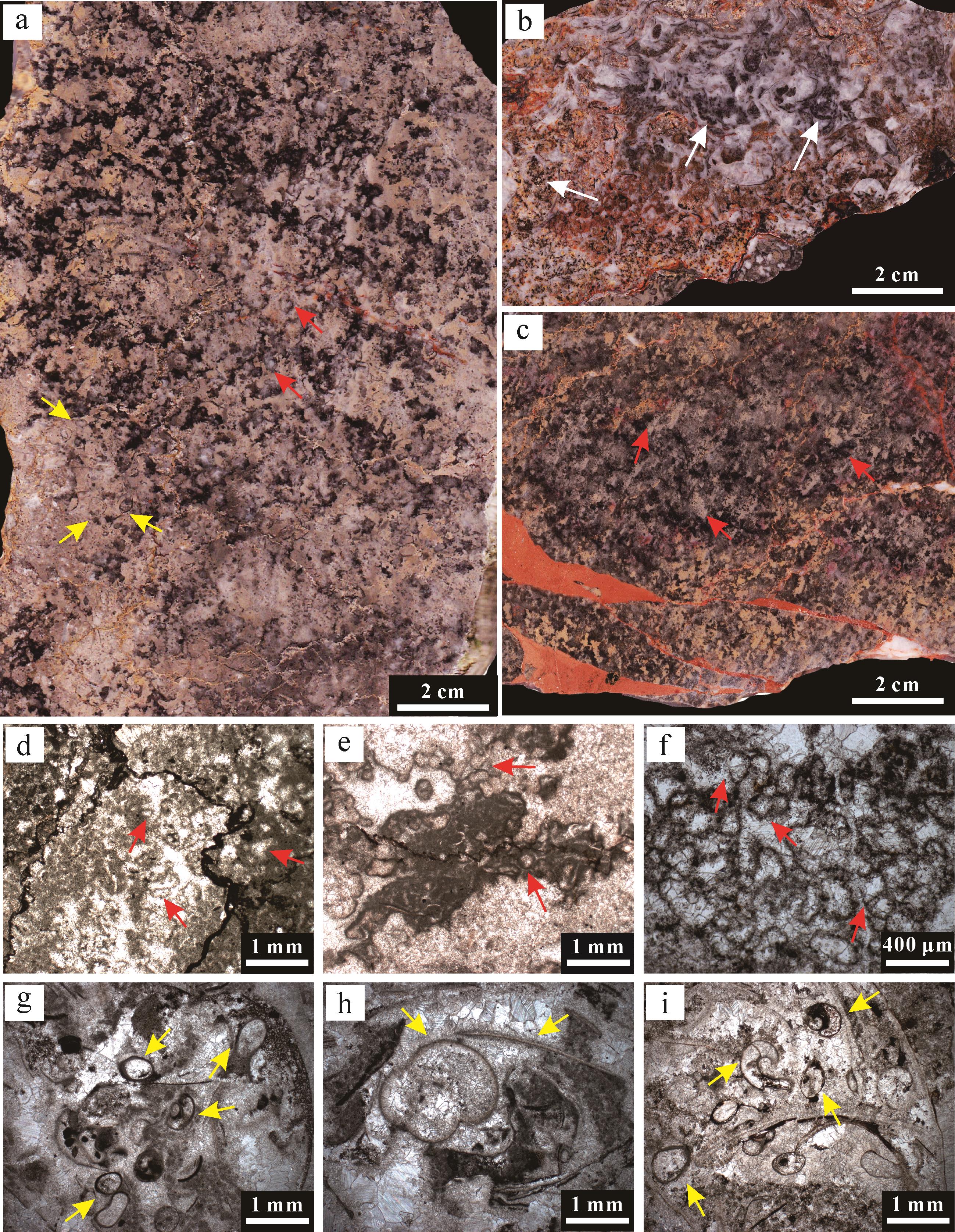
凝块石富含微生物化石,可鉴定为宽球藻目蓝细菌Gakhumella属(图4a~e)。Gakhumella也广布于华南地区其他剖面同时代的微生物岩序列中,但是相同的微生物化石被不同作者命名为不同微生物类群(王永标等,2005;Ezaki et al.,2008;Adachi et al.,2017;Fang et al.,2017;Wu et al.,2017,2022;Pei et al.,2019;Chen et al.,2022)。这种蓝细菌整体形态呈柱状,顶部为圆球状细胞(图4c),后方则保留6~25个杯状同心纹层(图4b,e),呈单体或簇状群体产出。Gakhumella在碳酸盐台地边缘环境中爆发,诱导碳酸盐快速沉淀,具有极高的钙化效率和碳酸盐生产效率,在微生物岩的形成过程中发挥重要作用(Wu et al.,2022)。

图 4 田湾剖面微生物岩中微生物和后生生物化石的显微特征
Figure 4. Microscopic features of microbial and metazoan fossils within microbialites at the Tianwan section
凝块石序列中夹多层富含双壳、介形虫、有孔虫、腹足和微管虫的生物介壳层透镜体(图3b,g~i、图4f,g)。微生物岩格架内也可见少量软体动物(双壳类与腹足类)和介形类化石碎片。这些生物碎屑与球粒、微晶基质共同构成颗粒支撑的泥粒灰岩组构。其中,双壳类壳体胶结物在显微镜下尤为显著。腹足类碎屑颗粒及壳体碎片也较为丰富,且常被微晶方解石包裹,指示了一种低能量的局限沉积环境。
-
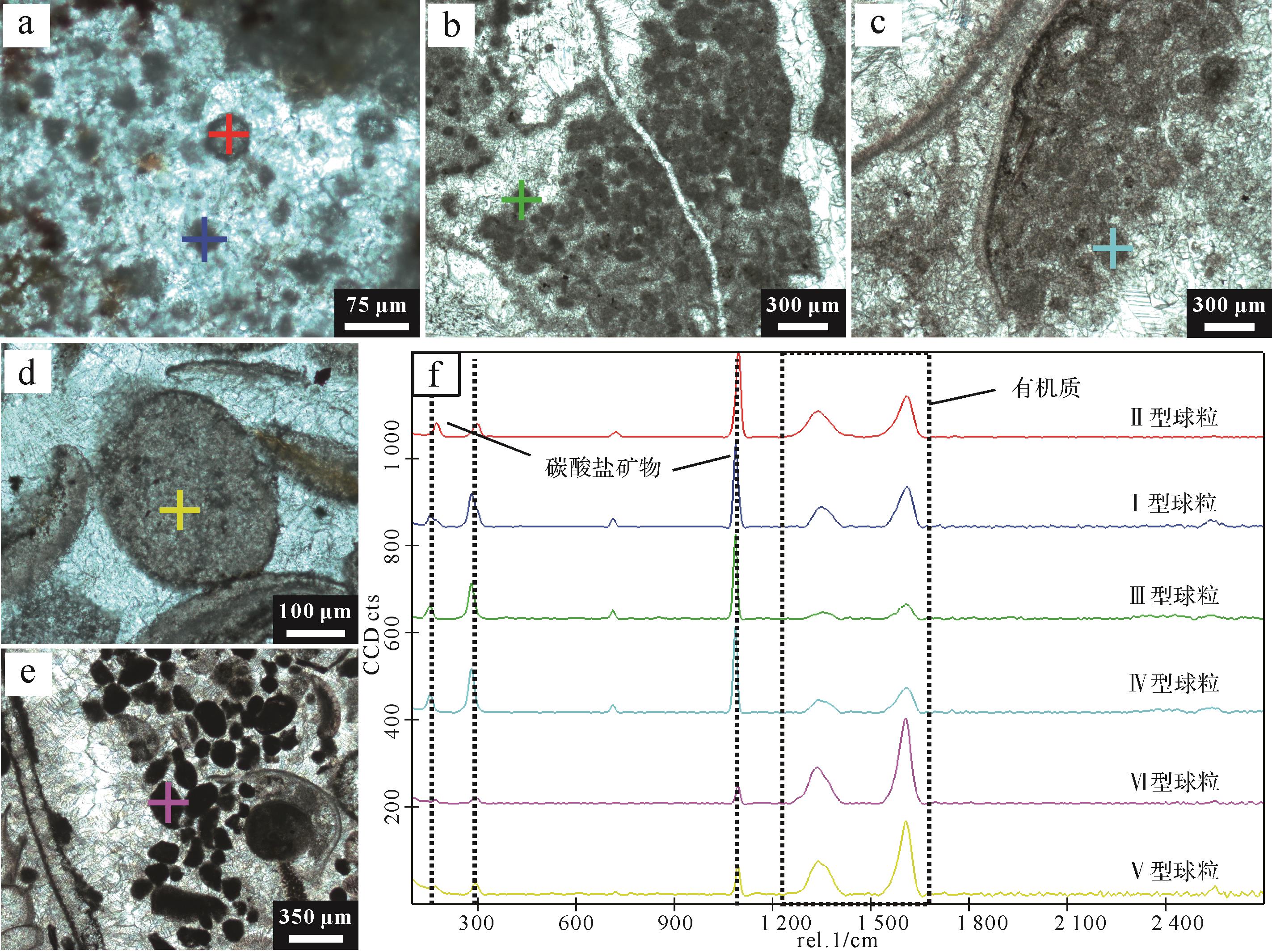
基于岩相学观察,并结合前人分类方案(Adachi et al.,2004),将赋存于微生物岩中的泥晶微球粒划分为六种类型:I型(原生微生物诱导钙化球粒,Microbially-Induced Peloid,MIP)、II型(重结晶微生物钙化球粒,Recrystallized Microbially-Induced Peloid,RMIP)、III型(陆源碎屑复合球粒,Aggregated Detrital Peloid,ADP)、IV型(骨骼间填充球粒,Intraskeletal Peloid,ISP)、V型(白云岩化泥晶球粒,Dolomitized Peloid,DP)和VI型(聚合型微生物诱导钙化球粒,Aggregated Microbially-Induced Peloid,AMIP)。各种类型泥晶微球粒的粒径大小、磨圆度、分选性、形态特征、矿物组成、重结晶程度及光谱和能谱特征简述如下(表2、图5~7)。
表 2 微生物岩中不同球粒的粒度、形态、矿物组成及光谱特征对比
Table 2. Comparisons of particle size, morphology, mineral composition and Raman spectral characteristics of various peloids within the microbialite
分类 I型球粒 II型球粒 III型球粒 IV型球粒 V型球粒 VI型球粒 特征 原生微生物诱导钙化球粒 重结晶微生物钙化球粒 陆源碎屑复合球粒 骨骼间填充球粒 白云岩化泥晶球粒 聚合型微生物诱导钙化球粒 粒度 细粒 细粒 中粒 中粒 粗粒 中粒 颗粒圆度 次圆状 次圆状 次棱角状 次圆状 棱角状 次棱角状、次圆状 分选性 良好 良好 一般 良好 一般 一般 基质矿物组成 亮晶方解石 亮晶方解石 泥晶方解石 泥晶方解石 亮晶白云石 亮晶白云石 胶结物类型 钙质泥晶胶结 钙质与少量白云质泥晶胶结 钙质泥晶胶结 钙质泥晶胶结 白云质泥晶胶结 白云质泥晶胶结 重结晶特征 无显著重结晶 核部重结晶显著,方解石为主 无显著重结晶 局部重结晶 局部重结晶 局部重结晶 分布位置 下部凝块石的方解石亮晶中 下部凝块石的方解石亮晶中 中下部凝块石的泥晶或亮晶区域 中部介壳层双壳化石内部/间隙 顶部介壳层生物化石碎片间隙 顶部介壳层生物化石腔体内部 颜色 黑色 黑色 棕色 黑色 深棕色 黑色 平均粒径/µm 45.3 50.5 151.5 125 291.4 175 粒径标准差/SD 15.6 7.3 53.6 31.6 114.3 52.1 粒径范围/µm 14~90 10~130 50~350 70~210 80~600 70~380 荧光特征 荧光响应弱 边缘荧光响应强 荧光响应弱 荧光响应弱 边缘荧光响应强,核部减弱 核部荧光响应强,边缘逐渐减弱 拉曼光谱特征 D1、G峰显著,可见D2、D4峰 D1、G峰清晰,可见方解石与白云石峰 D1、D2、G峰 D1、D2、G峰 D1、G峰显著,可见D2、D4峰,可见大量白云石峰 D1、D2、G峰显著,可见白云石峰 主要元素组成 富含Ca、O Ca和O含量最高,Mg含量略高 富含Ca、O,伴随Si、P、S少量富集 Ca和Si为主,少量Mg和Fe Mg/Ca显著升高,伴随Fe、S的富集 Mg/Ca显著升高,伴随P、Fe的富集 
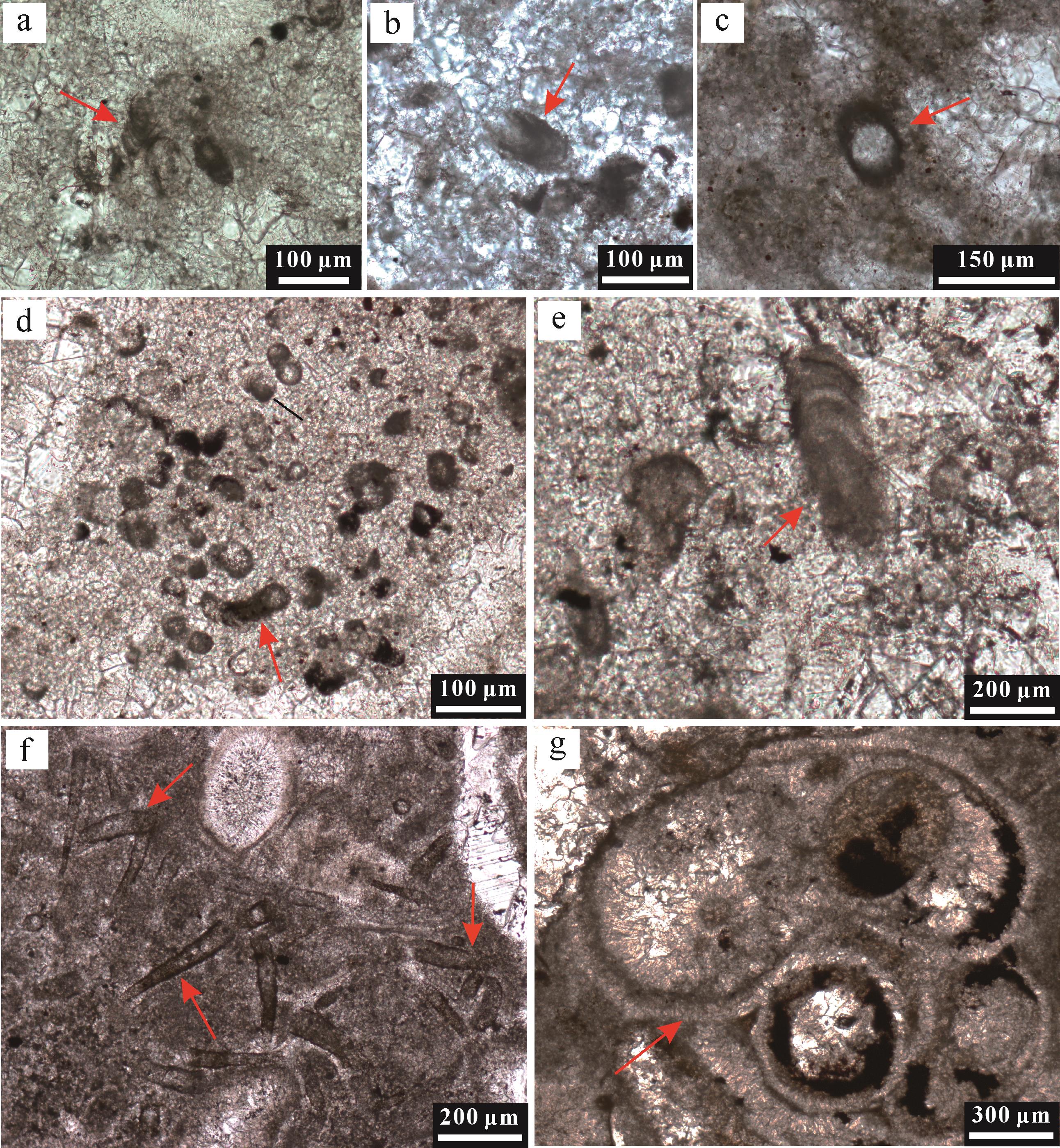
图 5 I~III型球粒的光学显微镜下特征
Figure 5. Photomicrographs of peloids I⁃III

图 6 IV~VI型球粒的光学显微镜下特征
Figure 6. Photomicrographs of peloids IV⁃VI

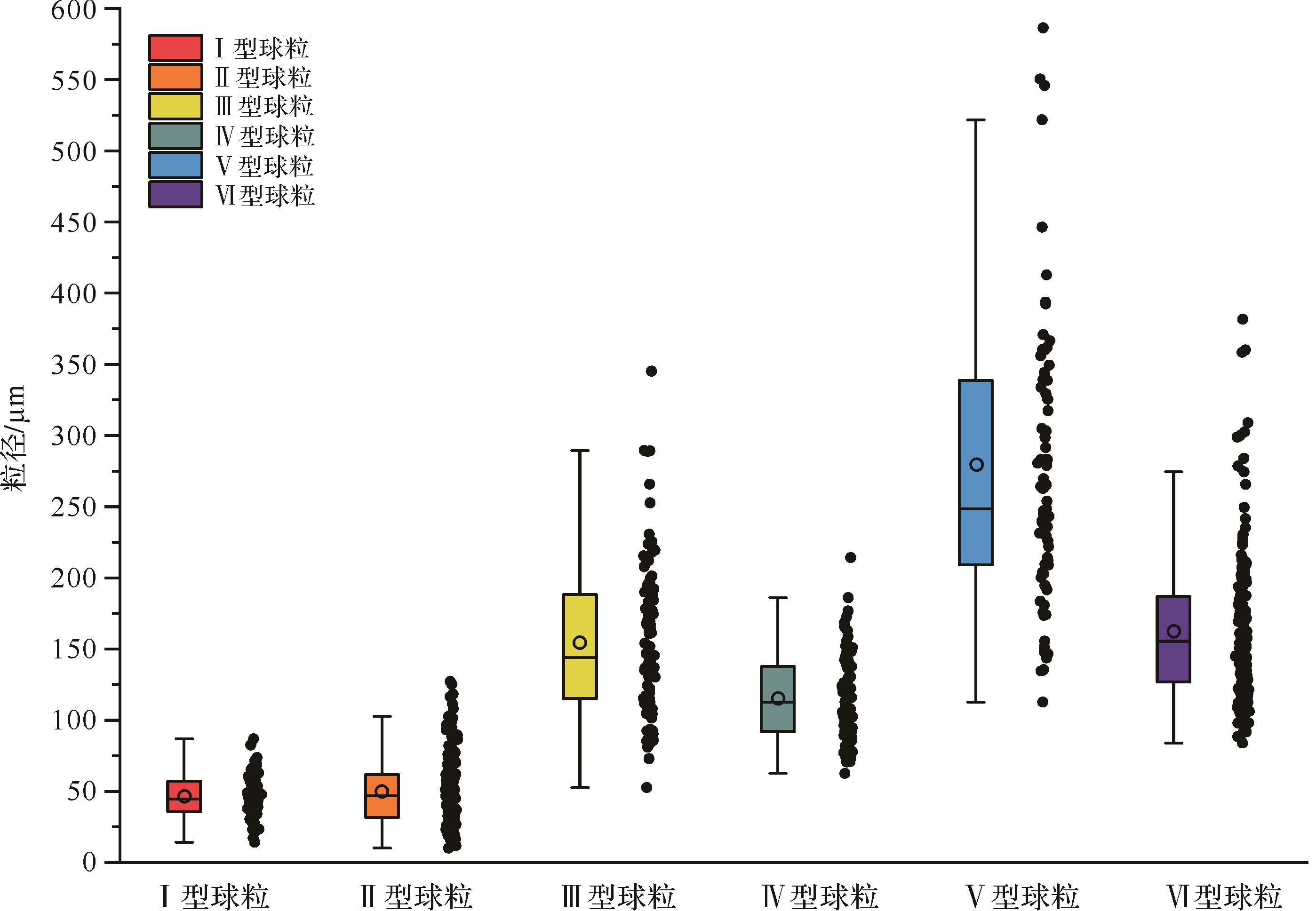
图 7 不同类型球粒结构的粒径统计结果对比
Figure 7. Comparison of particle size statistics of various peloids
I型球粒(MIP)产自微生物岩底部纹层状凝块石,赋存于亮晶方解石区域(图5a~e),呈黑色细小、次圆状颗粒,由泥晶方解石组成,基质为泥晶胶结(图5a,b),颗粒磨圆度一般,分选性较好。其粒径大小分布区间窄,介于14~90 µm,均值为45.30 µm,中位数为44.54 µm(图7)。
II型球粒(RMIP)也赋存于微生物岩底部纹层状凝块石的亮晶方解石区域(图5a~e),其颜色、形态特征、磨圆度、分选性和基质结构均与I型球粒相似,但有机质残留主要出现在球粒边缘,其核部被重结晶方解石和少量白云石填充(图5d,e)。其粒径大小介于10~130 µm,与I型球粒相近且范围略大,平均粒径为50.50 µm,中位数为47.00 µm(图7)。
III型球粒(ADP)产自微生物岩中下部的凝块石中,赋存于泥晶方解石区域(图5f~j),部分散落在方解石亮晶区域,与囊泡结构共存(图5h),呈棕色次棱角状(图5j)至椭球形,边缘比较弥散,具有模糊边界,分选性一般,其基质为泥晶胶结(图5f)。其粒径较前两类球粒明显大(图4f~i),其大小分布区间较宽,为50~350 µm,均值达到151.50 µm,中位数为144.24 µm(图7)。
IV型球粒(ISP)产自微生物岩中部的介壳层内,通常出现在双壳化石的介壳碎片附近,黑色次圆状形态,分选良好,基质为泥晶胶结(图6a,b),部分球粒内部被重结晶微晶方解石填充(图6c)。其粒径大小介于70~210 µm,均值约为125 µm,中位数为112.64 µm(图7)。
V型球粒(DP)和VI型球粒(AMIP)都分布于微生物岩顶部的介壳层中,球粒形态不规则,有机质呈絮状或丝状分布,部分重结晶显著(图6d~j)。V型球粒主要以一个破碎的介壳碎片为核心,周围被有机质包裹,总体结构与个体较小、发育不全的核形石颗粒非常类似,但其生物碎屑核心的泥晶化严重,外部包裹的有机质为白云质泥晶胶结,不发育任何纹层或圈层结构(图6e,f),因此与核形石有明显区别。其磨圆度和分选性都极差,粒径大小不均、分布范围宽,为80~600 µm,均值达291.40 µm,而中位数为248.25 µm(图7)。
VI型球粒也通常与生物化石碎屑共同出现,为次棱角状或次圆状形态,无核心结构,分选性较差,基质为泥晶胶结(图6g~j);其粒径大小介于70~380 µm,均值达175 µm,中位数为155.49 µm(图7)。
综上,I~III类球粒产自微生物岩的凝块石组构中;IV~VI类球粒则产自微生物岩的介壳透镜体夹层中。它们在颜色、形态、矿物组成和粒径分布上均存在明显差异。粒径统计表明,I型球粒的粒径大小分布区间最窄,其均值和中值都最小;II型球粒的粒径大小与I型类似,比后者稍大一点,但粒径分布标准差最小。相反,V型球粒粒径大小分布范围最宽,均值和中值也最大。总体上,球粒的粒径大小顺序大致为:V型>III型>VI型>IV型>II型>I型球粒(图7)。有机质通常残留于球粒的边缘或核部,呈现团簇状或絮状形态。总之,不同球粒的地层分布、形态特征和粒径大小受沉积环境的水动力水平、微生物活动强度和成岩作用的影响程度等因素控制。
-
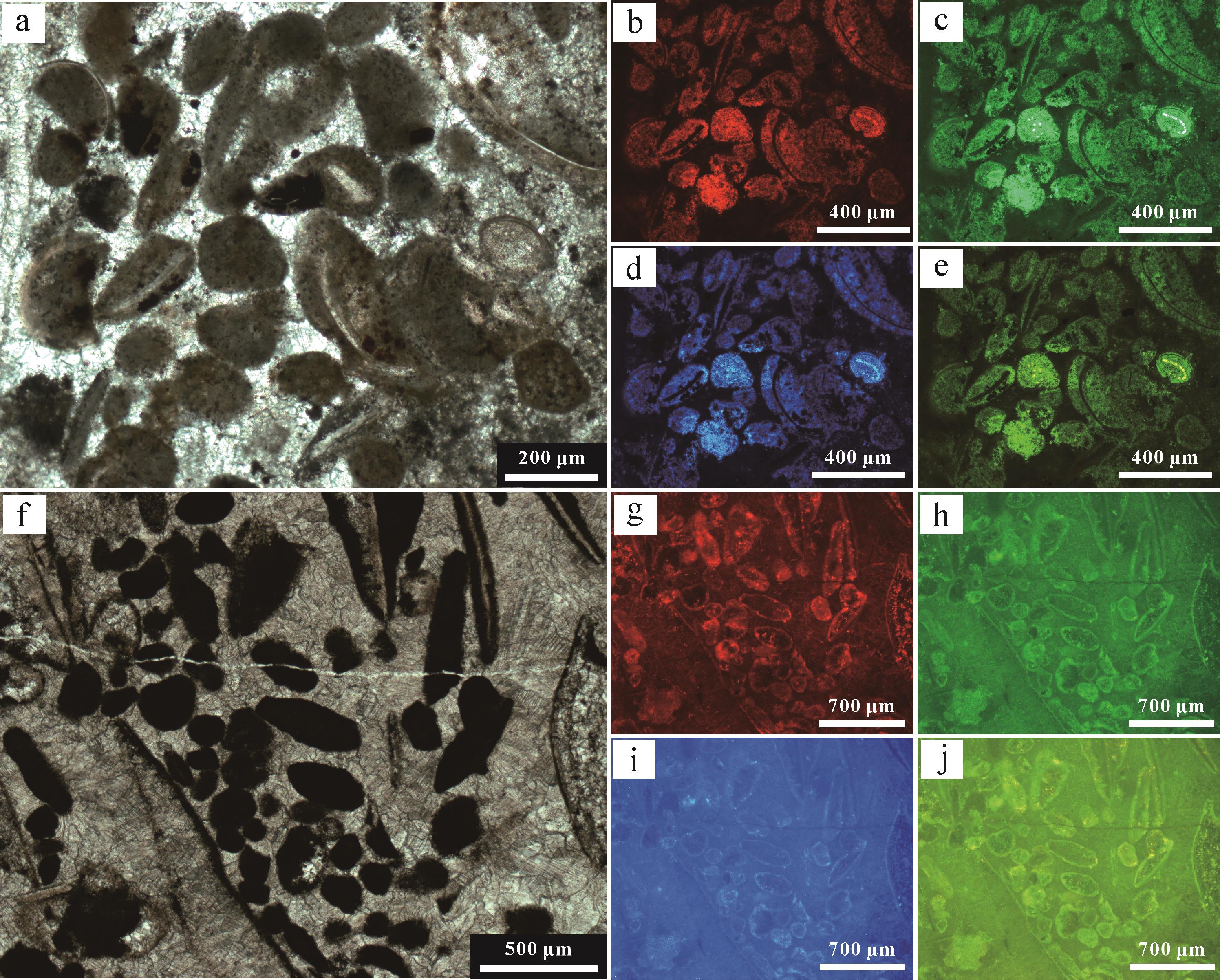
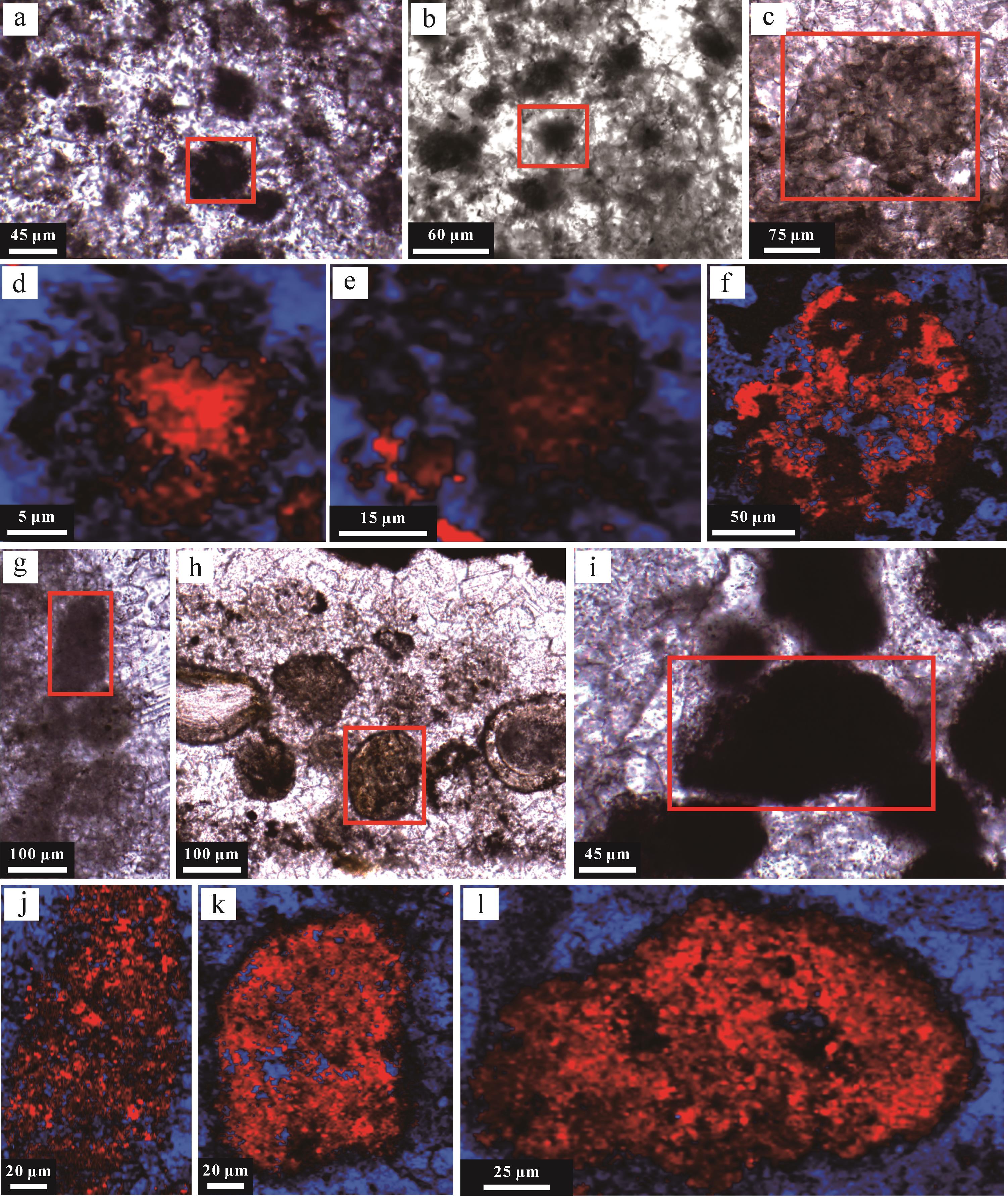
利用激发波长分别为430 nm(紫外)、365 nm(紫外)和450 nm(蓝色)的荧光,对有机团簇内有机物的分布进行了成像分析。其中,V型和VI型球粒表现出较为强烈的荧光响应(图8),其他类型球粒的荧光响应相对较弱,表明有机质降解程度存在差异。此外,V型球粒的残留有机质大多出现在球粒核部介壳碎片周围,呈现典型团块状聚集体(图8a~e);VI型球粒的残留有机质则多集中在球粒的边缘区域(图8f~j),指示有机质聚集或降解过程存在异质性。生物化石及其碎片周围的泥晶套均具有显著的荧光效应,指示大量的有机质。

图 8 不同激光照射下V和VI型球粒的荧光特征
Figure 8. Fluorescence photographs of peloids V and VI in different laser irradiation
-
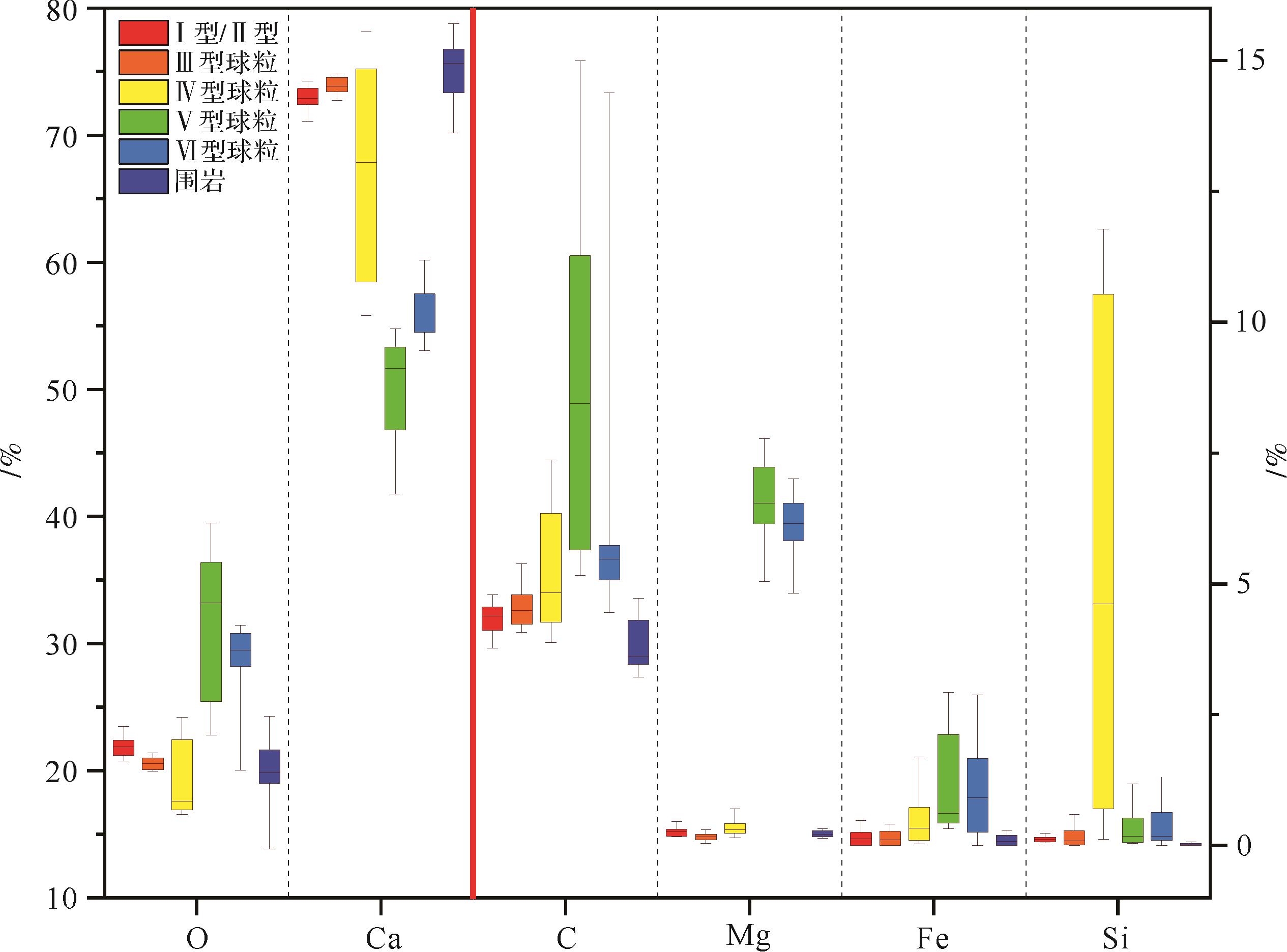
对各种球粒进行大量的能谱分析,获得大量的元素重量百分比数据后进行数理统计分析,计算出元素百分比的置信区间、中值和均值,并将其与围岩相同数据进行对比(图9)。其中,I型和II型球粒元素组成相似,分布层位相同,将它们两者的元素能谱分析结果合并表示。统计发现,不同类型球粒的主要组成元素均为Ca和O元素,且Ca的相对含量最高。有些球粒相对富集Si或Mg、Fe。球粒与围岩的K、P和S相对含量均低于0.8%,且无明显差异性。围岩则主要由Ca、C、O元素组成,几乎不含K、Mg、Fe、Si、P、S元素,其含量均不超过1%。I和III型球粒与围岩的成分近似,主要由碳酸钙(Ca、C、O)组成,其余元素占比均低于1%。IV型球粒中Si元素相对含量的占比高(>5%)。V和VI型球粒Mg相对含量的占比高(>6%),而Ca相对占比略低,指示白云岩化作用的特征。

图 9 不同球粒的能谱分析与元素组成对比
Figure 9. EDS analysis and elemental composition comparisons of various peloids
-
拉曼光谱分析表明,所有类型的球粒在1 080 cm-1处均出现强烈的方解石特征峰(图10),这是方解石C-O键对称伸缩振动的典型响应,指示碳酸盐矿物是球粒的主要组成成分。

图 10 不同类型有机质球粒结构的拉曼光谱点分析结果
Figure 10. Raman spectral point analysis results for different types of organic matter spheroid structures
部分球粒中还出现~1 090 cm-1附近的白云石特征峰,由白云石的C-O键伸缩振动而产生,与方解石峰的位置略有偏移(Dufresne et al.,2018)。白云石特征峰在部分球粒中表现明显,说明它们可能经历了不同程度的白云石化作用,与岩石薄片中观察到的重结晶特征和白云质胶结物的出现相符。此外,在1 350 cm-1(D峰)和~1 610 cm-1(G峰)处均检测到显著的有机质特征峰(王民和Li,2016),分别对应于无序碳的缺陷模态振动和石墨化碳的平面振动,指示球粒中有机质普遍存在(图10)。
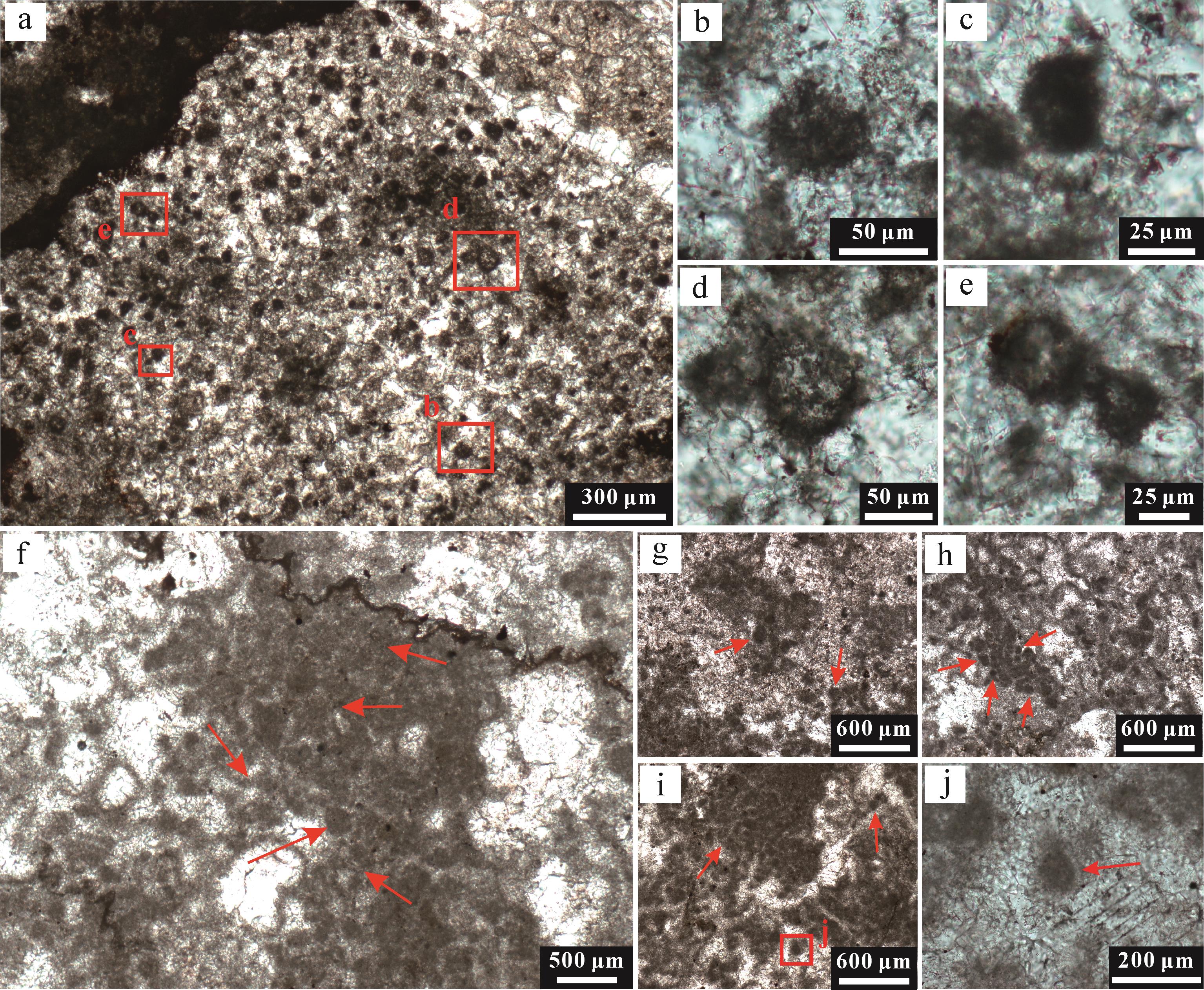
拉曼光谱面扫结果进一步揭示(图11),有机质主要赋存于球粒或者生物化石和介壳碎片外部的泥晶套中,且在围岩中的含量显著降低。此外,球粒内部的有机质多呈团簇状或絮状分布,特别是在I、V和VI型球粒中保存较为完好,可见有机团块的集中分布(图11d,k,l)。其他类型的颗粒受成岩作用影响较显著,在重结晶和成岩烘烤作用的影响下,有机质小规模团簇状聚集,整体呈絮状分布(图11f,j),部分球粒内部有机质被大量降解,主要聚集在颗粒边缘(图11e)。

图 11 不同类型有机质球粒结构的拉曼光谱面分析结果
Figure 11. Raman spectral mapping of various peloids
-
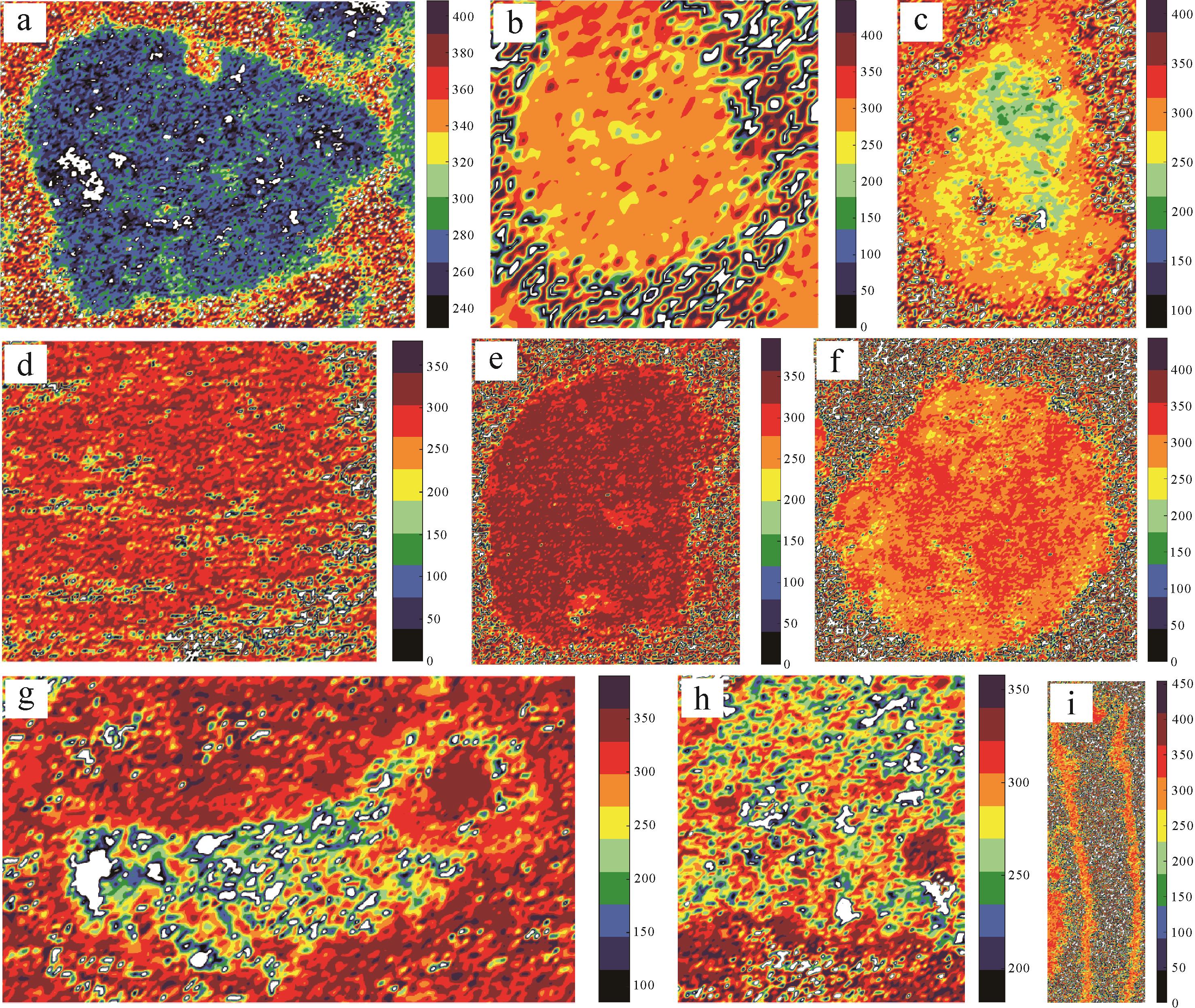
由于温度对拉曼光谱结构影响是不可逆的,因此,研究剖面的蓝细菌Gakhumella化石、有孔虫化石、不同类型球粒以及介壳碎片外部泥晶套的有机质光谱信息,可恢复成岩过程中有机质受到加热烘烤程度的影响程度,并以峰期成岩温度差异的形式体现(图12)。

图 12 研究剖面的球粒、生物化石及介壳碎片外部泥晶套的拉曼温度平面分布图
Figure 12. Raman temperature maps for various peloids, fossils and micritic envelopes on the outside of shell fragments from the study section
这些物质的拉曼拟合曲线在1 100~1 700 cm-1段均出现有机质峰形。通过洛伦兹拟合发现,有机质含量高的球粒(II,III,IV型)和有孔虫化石含有D1、D2和G峰;而I、V和VI型球粒,Gakhumella及介壳碎片外部泥晶套均含有D1、D2、D4和G峰。通过拟合后的频谱峰形特征对比,计算其成岩烘烤温度(表3)。同时,基于不同的地质温度,评价有机质在成岩作用过程受成岩烘烤程度(表3)。此外,温度面扫技术可揭示不同温度的有机质在球粒、生物化石和围岩中的分布特征(图12)。
表 3 利用拉曼温度计对不同球粒、生物化石和介壳碎片泥晶套的有机质成岩烘烤温度计算结果
Table 3. Calculated peak period alternation temperatures of various peloids, fossils and micritic envelopes of shell fragments using Raman thermometer
类型 温度范围/℃ 集中分布范围/℃ 成岩烘烤程度 物质来源相似性 I型球粒 公式1:240~340公式2:310~380公式3:365~445 公式1:240~300公式2:310~330公式3:380~410 成岩烘烤程度最低 与VI型球粒和泥晶套接近 II型球粒 公式1:220~310公式2:210~430公式3:0~256 公式1:270~310公式2:210~320 公式3:64~256 成岩烘烤程度不均,核部重结晶严重,温度差异显著 与I型球粒有一定演化关系,但变质程度较高 III型球粒 公式1:260~340公式2:260~370公式3:330~500 公式1:260~340公式2:280~340公式3:400~460 高成岩烘烤程度,在公式2和公式3计算结果中尤为显著 与V型球粒物质来源最为接近 IV型球粒 公式1:220~350公式2:210~370公式3:350~470 公式1:220~280公式2:270~320公式3:390~430 公式1、公式2成岩烘烤程度较低,公式3中成岩烘烤程度中等 与Gakhumella和泥晶套接近 V型球粒 公式1:320~360公式2:250~380公式3:380~475 公式1:320~360公式2:286~350公式3:410~445 最高成岩烘烤程度,与III型球粒热演化规律一致 III型球粒和V型球粒的部分物质来源极为接近 VI型球粒 公式1:170~310公式2:250~370公式3:350~455 公式1:180~248公式2:260~330公式3:360~390 成岩烘烤程度最低,可能为I型球粒的演化产物 与I型球粒和泥晶套相似 Gakhumella 公式1:170~410公式2:255~340公式3:360~510 公式1:300~360公式2:280~330公式3:400~460 公式1和公式2成岩烘烤程度中等,公式3显示较高程度成岩烘烤 与后生生物化石接近 后生生物化石 公式1:220~385公式2:255~340公式3:330~465 公式1:270~360公式2:280~330公式3:425~455 公式1和公式2成岩烘烤程度中等,公式3显示较高程度成岩烘烤 与泥晶套成岩烘烤规律相近 泥晶套 公式1:250~320公式2:310~350公式3:380~435 公式1:250~320公式2:320~340公式3:400~425 低至中等成岩烘烤作用 与VI型球粒最为相似 注: 公式1与公式2计算结果相近,均表现为较低的成岩烘烤程度,与峰形拟合结果统一,公式3计算得到的温度范围与实际峰形所处温度范围差异较大,故主要参考公式1、2特征总结规律。地质温度计算表明,I型球粒(MIP)成岩烘烤温度最低,其内部有机质的成岩烘烤温度主要介于260 ℃~280 ℃,少量有机质的成岩烘烤温度低至240 ℃或高达300 ℃。球粒内部和围岩中的有机质成岩烘烤温度对比强烈(图12a)。II型球粒(RMIP)的核部重结晶严重,温度主要集中在270 ℃~310 ℃,与围岩的有机质烘烤温度相近(图12b),边部有机质的成岩烘烤温度介于350 ℃~400 ℃。由此可见,II型球粒的有机质经历了一定程度的成岩烘烤作用改造,但核部因重结晶作用,局部成岩烘烤程度较为突出,温度分布存在一定的不均性。
VI型球粒(AMIP)的成岩烘烤温度范围较宽,介于170 ℃~310 ℃,其中核部温度主要集中在180 ℃~248 ℃,可见明显的低温核部区域,边部温度则相对较高,且明显高于围岩有机温度。此温度特征显示出其中等成岩烘烤程度,核部的低温保护区可能是由于包裹效应所致,可能是I型球粒被海水有机物包裹的产物,其成岩烘烤温度也与介壳碎片外部泥晶套的温度变化情况较为相近,略低于生物化石(图12c)。
III型球粒(ADP)的成岩烘烤温度范围为260 ℃~340 ℃(图12d),V型球粒(DP)为320 ℃~360 ℃(图12e),反映了这两类球粒在成岩过程中承受了较高的成岩烘烤程度,表明部分有机质受到了更强烈的成岩烘烤,具有较高的热演化影响。IV型球粒(ISP)的成岩烘烤温度范围为220 ℃~350 ℃,虽然成岩烘烤温度分布范围较宽,但核部有机质集中分布区域的成岩烘烤温度相对较低(220 ℃~280 ℃),属于中低等程度成岩烘烤;分布于球粒边缘部分的有机质成岩烘烤温度相对较高(图12f)。其温度变化情况与蓝细菌Gakhumella的成岩烘烤温度最为接近。总体上,球粒边缘部分的有机质受成岩烘烤的温度明显高于围岩的有机质。
蓝细菌Gakhumella的成岩烘烤温度范围为300 ℃~360 ℃(图12g),后生生物化石(如有孔虫)介于270 ℃~360 ℃(图12i)。有机质主要分布于化石的纹层及边缘部位,各纹层内部有机质几乎被完全分解。介壳碎片外部泥晶套包裹物的成岩烘烤温度范围为250 ℃~320 ℃(图12h)。
-
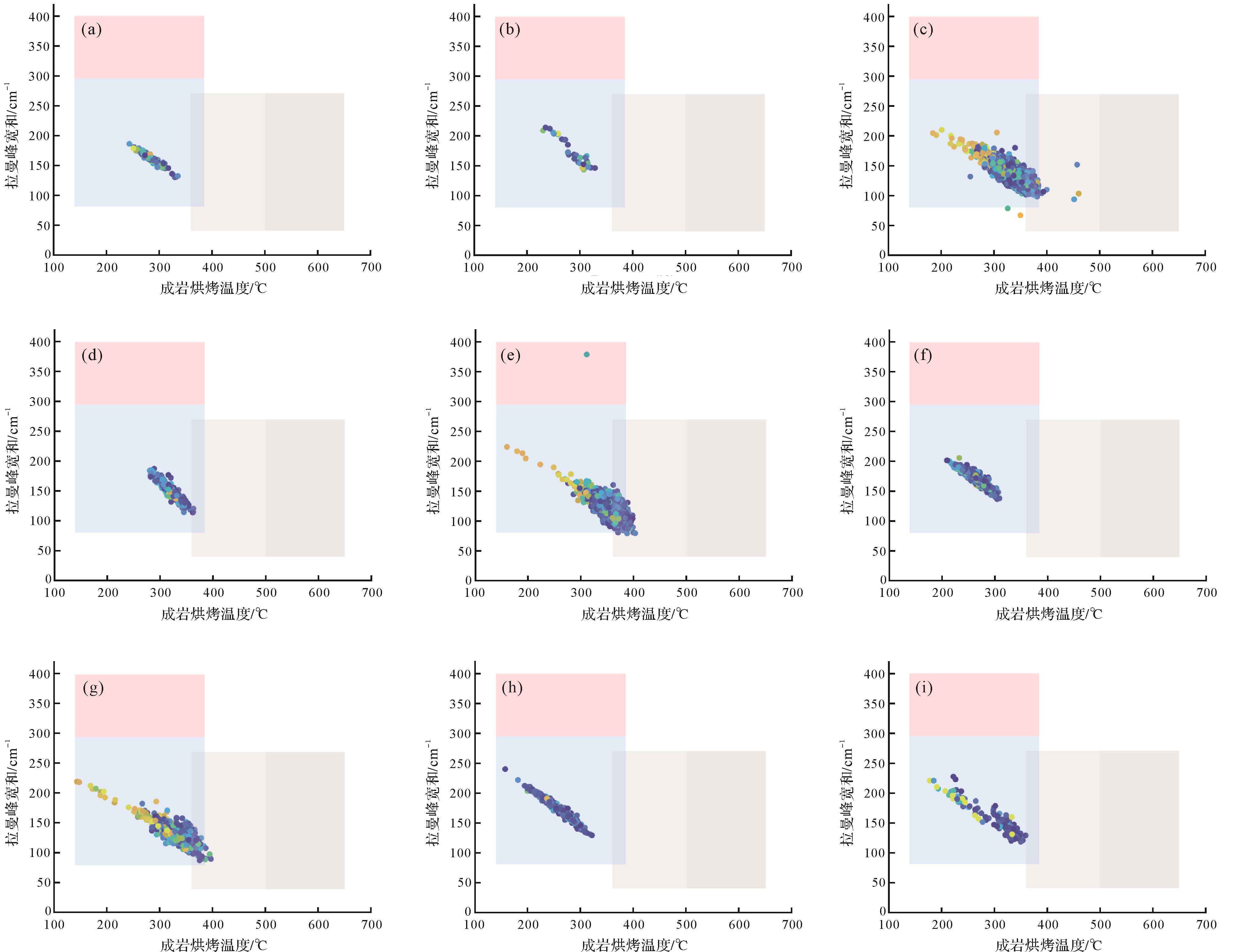
在成岩烘烤作用的影响下,球粒中的有机质往往被降解,受到烘烤作用的影响,有机质渐进式裂解,H、O、N等元素被排出,其碳原子排布方式逐渐由无序态向石墨化态转变(Canfield et al.,2021),该过程形成的一系列材料被称为碳质物质(Carbonaceous Matters, CM),变质温度升高的同时,石墨化程度也不断提高,最终形成堆叠的石墨烯薄片(Sparkes et al.,2013,2020)。Sparkes et al.(2020)根据碳质材料的拉曼光谱结果,将拉曼峰宽和与成岩烘烤温度计算结果作为参数,表征有机质的石墨化程度,分为四个等级,即无序态,初级石墨化,中级石墨化与高级石墨化。
研究剖面的有机质整体处于初级石墨化阶段,但即便在同一种微生物岩中,不同位置的有机质也展示了从低温到高温的广泛温度特征,有机质在成岩作用下经历了复杂的成岩烘烤过程,可能受包括有机质的原始来源、沉积环境、成岩流体及局部埋藏环境等多种因素的影响。基于拉曼温度计算结果中R²大于0.97的数据点进行拟合(图13),可以观察到不同类型球粒及微生物—生物化石在石墨化程度上的变化趋势,揭示了它们在成岩烘烤作用下的演化过程。

图 13 不同类型球粒、生物化石和介壳碎片外部泥晶套的有机质石墨化程度分析
Figure 13. Graphitization degrees of organic matter preserved in various peloids, fossils and envelopes at the outside of the shell fragments
在I型球粒(MIP)中,大部分有机质处于初级石墨化状态,但仍有部分区域保留无序化结构,表明其经历的成岩烘烤作用较弱。这与其形成初期的埋藏环境较浅或局部成岩流体的影响有关,部分有机质未经历充分热演化影响(图13a)。II型球粒(RMIP)的有机质保存较差,主要表现为局部重结晶方解石填充,仅边部保留较强的有机信号。其成岩烘烤程度略高于I型球粒(图13b)。VI型球粒(AMIP)在温度与峰宽上的变化范围几乎与I型和II型球粒重合,表明其有机质的石墨化程度与这两类球粒相似。这种相似性与其形成机制有关,VI型球粒可能是I型球粒被其他有机碎屑颗粒包裹的产物,并经历了类似的成岩过程。因此,VI型与I型球粒类似,它们在成岩演化过程中均未表现出较高的成岩烘烤程度(图13f)。
I、II型球粒来自微生物岩底部的凝块石,其有机质可能来源于当时爆发的微生物,或滞留在海水中的有机质。值得注意的是,微生物岩底部与下伏二叠系生物碎屑灰岩之间出现平行不整合,说明后者经历过暴露,而微生物岩则是大灭绝事件后华南发生区域大海侵时期的沉积产物(王永标等,2005;刘建波等,2007;Chen et al.,2022),因此,微生物岩底部球粒的有机质更有可能来源于下伏地层风化产物。然而,I、II型球粒有机质的成岩烘烤温度明显低于二叠系生物碎屑灰岩中有机质的平均温度,表明有机质来源既不是下伏二叠系风化产物,也不是海水滞留有机质,后者主要来自暴露出海平面的早期地层,经历大陆风化后被搬运至海洋,从而滞留在海水中。由此可见,这些有机质形成的最初时间早于其再沉积时间,因而相较于微生物岩原生有机质经历更多次成岩烘烤,其温度也显著更高(Canfield et al.,2021)。相反,I、II和VI型微球粒有机质的成岩烘烤温度与微生物岩中Gakhumella和后生生物化石的温度大体一致,且稍稍低于后者,说明这些有机质极可能来源于当时爆发的蓝细菌的生物分解或代谢活动。这些有机质埋藏之后,往往比Gakhumella和后生生物化石的有机质保护得稍好,因此其成岩烘烤温度也略低(Canfield et al.,2021)。
III型球粒(ADP)和V型球粒(DP)的有机质表现出最高的成岩烘烤温度,与I、II、VI型球粒的有机质存在明显差异。III型球粒的少部分区域已达到中级石墨化水平(图13c),拉曼峰宽较窄,表明其有机质结构更有序,成熟度较高(王民和Li,2016)。同样,V型球粒的成岩烘烤温度和峰宽分布较为离散,但整体上表现出较高的有机质结构有序度和成熟度(图13e)。III和V型球粒有机质的成岩烘烤温度明显较Gakhumella和后生生物化石的温度高。后者代表微生物岩原生有机质的成岩烘烤温度,说明III和V型球粒有机质可能承受了比微生物岩原生有机质更多次数的成岩烘烤作用。正如前文所述,该类有机质可能来源于长期滞留在海水中的有机质。这是因为早三叠世华南陆缘构造活动活跃,风化作用增强导致陆源物质输入增加,这些有机质最初来自陆地环境的页岩或泥质沉积物(杜远生等,2013),在其源岩成岩时,这些有机质就已经经历至少一次成岩烘烤作用,然后,经过暴露风化作用和沉积过程,汇聚到海洋之中。最后,通过聚合黏结的方式形成V型球粒的重要成分。微生物岩成岩后,这两类有机质再次受到成岩烘烤,导致它们的温度明显较同期原生有机质(如Gakhumella)的烘烤温度要高(图13c,e)。
IV型球粒(ISP)的成岩烘烤特征与Gakhumella和有孔虫化石较为接近(图13d,g,i),表明其有机质主要来源于微生物岩内部微生物的有机质分解或代谢活动的产物。同时,IV型球粒主要填充于生物骨架内部,因此其有机质在埋藏过程中受到一定的保护,导致其有机质成岩烘烤温度相对较低,发育与微生物化石相似的热演化特征(Reid,1987;刘贝,2023)。此外,生物介壳层中大量的生物碎屑在埋藏过程中起到保护作用,使得球粒表现出渗流沉淀的特点,其中有机质也因此免受降解。
Gakhumella的有机质在其化石纹层边缘的保存效果较好(图13g)。这种特征表明微生物在矿化过程中,局部纹层结构能够在一定程度上保护有机质,使其在成岩过程中受到成岩烘烤的影响较弱。有孔虫壳体结构较为坚固,有助于在埋藏和成岩过程中对有机质进行保护(图13i)。这两类有机质代表了微生物岩形成过程中的原生有机质,它们的成岩烘烤温度代表了微生物岩成岩过程中承受的成岩烘烤程度。
此外,一些介壳碎片周围发育的泥晶套也显示出较高的有机质富集特征,其成岩烘烤温度表明泥晶套整体仍处于初级石墨化阶段,说明介壳碎片泥晶套的有机质主要来自微生物岩系统内部,且与微生物的呼吸作用密切相关。泥晶套有机质与微生物和后生生物的有机质相同,仅经历了一次成岩烘烤(图13h),因此成岩烘烤温度相对较低。大量有机质在介壳碎片外部泥晶套的富集也说明,微生物岩内部的有机质主要通过泥晶化的方式进行保存(Sun and Wright,1989;Neuweiler,1993),而后期成岩过程中局部热液的影响可能导致有机质的进一步演化。
综上,微生物岩内部大部分泥晶微球粒的形成可能与有机质泥晶化作用有关,其有机质来源于原生微生物及后生生物化石,且由于重结晶或多期次有机质包裹保护的影响,残留的有机质受到成岩烘烤的程度有所区别。总体上,田湾微生物岩中大多数有机质(如I、II、IV、VI型球粒的有机质)在经历成岩烘烤作用后仍主要处于初级石墨化阶段,说明这些有机质来自微生物岩系统内部,主要由大灭绝后微生物爆发时期所产生,仅承受一次成岩烘烤作用影响。相反,III和V型球粒的有机质成岩烘烤程度较高,其部分有机质可能源自长期滞留在海水中的陆源有机物(Steinhauff et al.,2021)。因此,III和V型球粒的有机质可能经历了不止一次的成岩烘烤,其温度明显高于其他类型球粒的有机质。因此,拉曼温度计是判别地质历史时期有机质来源的重要手段。
-
田湾剖面微生物岩发育在PTME之后南盘江盆地孤立碳酸盐台地边缘,其沉积环境具有亚氧化、较低能的特点(刘建波等,2007)。亚氧化的水体环境有利于有机物的埋藏,而水动力条件的差异对球粒的形态与分选性起到关键的控制作用。泥晶球粒广布于微生物岩格架内部及生物介壳层透镜体内,且没有经历异地搬运,在原地形成和埋藏。泥晶球粒通常为颗粒小、球形至椭球形的隐晶质碳酸盐岩聚集体,缺乏明显同心纹层或圈层构造(Samankassou et al.,2005)。Adachi et al.(2004)首次描述了该剖面微生物岩的岩石组构和泥晶球粒特征,他们认为球状微生物死亡后,其躯体部分有机质分解并钙化,球状微生物内部被方解石填充,轮廓保存并形成球粒。对这些球粒细化观察发现,几乎没有球粒是微生物本身因有机质分解而钙化,并被方解石充填而形成。相反,大多数球粒是微生物活动与环境条件共同作用的产物。大灭绝之后,华南地区发生大规模海侵,导致微生物岩底部在不整合面之上再度发生沉积作用(王永标等,2005;刘建波等,2007;Chen et al.,2022)。由于微生物岩发育的地区为碳酸盐台地,这里陆源物质输入缺乏,微生物(特别是蓝细菌)非常发育,有利于碳酸盐沉淀。此外,由于微生物岩建造以凝块石为主,而后者通常是在相对低能环境中沉积(Ezaki et al.,2008;Kershaw et al.,2012),因此微生物岩的泥晶微球粒是原地沉积和埋藏的。
I、II型球粒出现在微生物岩底部纹层状凝块石中,其粒径较小、分选良好,指示相对高能环境,这与微生物岩发育初期在华南地区发生区域性的大海侵相一致(王永标等,2005;刘建波等,2007;Chen et al.,2022)。这些球粒中的元素组成以Ca、O为主,Si、Mg含量小于1%,有机质温度稍低于蓝细菌Gakhumella、后生生物化石的温度,并明显低于下伏二叠系亮晶生物碎屑灰岩中有机质的平均温度。此外,微生物岩底部未见以下伏灰岩为源岩的砾石,或者再沉积的产物,表明I、II型球粒的有机质主要来源于大灭绝后与海侵同步的微生物爆发时期原地微生物遗体或其代谢活动的产物,而非陆源物质因风化作用带入海水中的有机质。此时,微生物通过光合作用固定CO2,分泌胞外聚合物(EPS)形成黏性基质,以固定微生物群落、捕获沉积物,EPS中的多糖和蛋白质也可以作为成核位点,结合Ca²⁺和Mg²⁺并诱导碳酸钙沉积,促进方解石或白云石沉淀(田野和田云涛,2020),形成富含有机质的微生物席结构(Monty,1976;Chafetz,1986;Wu et al.,2022)。受到海侵初期较为高能的水动力条件的影响,微生物活动形成藻席后被扰动而破碎,进一步磨圆形成球粒结构。
III型球粒保存于微生物岩建造中下部的凝块石内,其形成时水动力条件相对较弱,相较于I、II型球粒有更长时间接受有机质沉积,表明III型球粒的有机质可能来自微生物的代谢产物或者滞留在海水中的有机质。然而,III型球粒的成岩烘烤温度高于蓝细菌Gakhumella和后生生物化石的温度,说明球粒中大部分有机质可能来源于滞留在海水中的有机质。后者主要来源于陆地风化产物,经过搬运作用被带入海洋并滞留在海水中,相较于微生物岩原生有机质,往往经历更多次成岩烘烤,因此成岩温度也显著更高(Canfield et al.,2021)。
IV型球粒保存于透镜状的生物介壳层内部,磨圆、分选良好,形成于相对高能环境,该类球粒具有模糊边界,成岩烘烤温度与Gakhumella接近,且其中Si元素显著富集(>5%),说明有机质主要来源于Gakhumella等微生物的躯体分解或代谢活动的产物,同时,微生物可能分泌大量EPS,捕获水体中黏土矿物等硅质碎屑,最终混合形成泥晶球粒(Hong et al.,2017)。
V型球粒粒径较大、分选较差,形态为棱角状,指示其形成时的水动力条件较弱,同层位部分生物碎屑被泥晶颗粒包裹形成类似核形石结构,表明该类球粒的形成可能与生物内碎屑的泥晶化密切相关(Flügel,2004)。该类球粒的有机物温度最高,受到成岩烘烤程度最强,表明其有机质多为海水滞留有机质。同时,可见拉曼白云石特征峰(1 090 cm-1),Mg/Ca也显著升高(>6%),表明白云岩化作用对其中有机质的改造程度较高。
VI型球粒保存于微生物岩顶部的生物介壳层中,形成于较高能动荡的微环境,其成因与生物碎片的泥晶化作用存在一定关联,是泥晶颗粒包裹生物碎屑与I型球粒类似物的产物,有机质主要来源于微生物岩原生的微生物躯体有机质分解或代谢活动的产物。
-
二叠纪末大灭绝后,微生物在浅水碳酸盐台地繁盛,特别是蓝细菌大量爆发(Derrien et al.,2023),它们通过光合作用、化能合成作用等途径,将无机碳(如CO2、HCO
在埋藏过程中,成岩作用引发的温度和压力变化使得有机质经历了成岩烘烤作用,导致其结构发生变化(Beyssac et al.,2002;Kouketsu et al.,2014;刘贝,2023)。特别是在较深的埋藏环境中,随着温度升高,有机质的分解速率加快,许多有机物质最终被转化为CO2或其他无机物,只有极少数的有机质通过矿化作用被封存,成为长期保存的深层碳。然而降解过程是不可逆的,随着成岩作用的进一步加深,有机质逐渐被分解和转化,最终导致碳的深层储存效率较低。
此外,浅水环境中的微生物大量爆发,其生成的有机质可能发生埋藏转移,如被运移到更深水的地区埋藏。然而,在华南地区PTME的深水相剖面,如鄂西地区的大峡口剖面,与微生物岩等时的深水相沉积物记录的TOC含量也非常低(Shen et al.,2015;Pei et al.,2021),说明当时华南海洋中并没有发生有机质埋藏转移现象。
综上,尽管大灭绝后火山作用刺激微生物爆发,并生成了大量的有机质,但生物降解、水动力作用以及成岩过程等因素共同作用,限制了其在沉积物中的埋藏效率,使得微生物岩中TOC含量普遍较低(Zheng et al.,2024)。微生物岩中的碳埋藏机制为我们理解古环境的变化和碳循环的演变提供了重要的地质证据,尤其是对于研究气候变化和地球历史时期碳循环的演化具有重要意义。
-
(1) 研究剖面微生物岩中球粒的形成主要与微生物活动密切相关,尤其是蓝细菌通过光合作用生成大量有机质,同时固定CO2,并通过分泌胞外聚合物(EPS)促进碳酸盐沉淀。球粒主要由有机质组成,形成过程中受沉积环境的水动力条件的影响。
(2) 根据形态特征和组成成分的不同,微生物岩的球粒可分为六种类型。其中,I和II型球粒的形成与微生物代谢作用有关;III型球粒的形成可能与海水中滞留有机质的富集以及新生变形作用相关;IV型球粒是微生物有机质与黏土矿物的混合物;V型球粒的形成可能与生物内碎屑的泥晶化作用密切相关,多来源于海水滞留有机质富集。VI型球粒是生物碎屑泥晶化的产物,其有机质主要来源于原生微生物有机质。
(3) 拉曼光谱地质温度计分析显示不同类型球粒的有机质经历了不同程度的成岩烘烤作用。大多数球粒有机质受成岩烘烤作用影响较小,处于初级石墨化阶段;III和V型球粒则保存有受到多次成岩烘烤作用影响的高温有机质,达到中级石墨化程度,可能源自滞留在海水中的陆源有机质。这表明拉曼地质温度计分析是检验有机质热演化、判别有机质物质来源的重要技术工具。
(4) 尽管微生物岩中微生物爆发生成大量的有机质,但频繁变化的氧化—还原状态和急剧升高的海水温度加剧了大量溶解有机质被氧化为CO2释放到大气中。仅有少量溶解有机质变成POC,并通过沉积被封存,成为由有机质内碎屑组成的球粒结构。颗粒形成后,成岩作用的成岩烘烤分解了大量有机碳,只有极少量的有机质矿化被封存,从而解释了微生物岩体系TOC含量普遍偏低的现象。
Classification and Organic Provenance of Microbialite Peloids in the Early Triassic
-
摘要: 目的 二叠纪末大灭绝后,微生物岩广布于浅海台地环境。该体系中微生物爆发与古生产力低下的矛盾长期以来备受关注。解剖贵州罗甸地区田湾剖面三叠系底部的微生物岩建造,关注泥晶球粒的岩石学特征及其有机质热演化和物质来源,厘清微生物成因的有机质和海水滞留有机质对球粒形成的影响,可为探究大灭绝后微生物岩体系中碳循环机制提供实证。 方法 采用光学显微镜、荧光、扫描电镜(SEM)与能谱(EDS),以及激光拉曼光谱分析等原位微区分析技术,对赋存于田湾剖面微生物岩中多种类型的泥晶球粒进行分析。 结果 泥晶球粒可分为六种类型。其中,I、II和VI型球粒的有机质成岩烘烤温度低,主要来源于微生物岩系统内部原生蓝细菌和其他微生物的光合作用和代谢产物。VI型球粒是泥晶颗粒包裹I型球粒的产物,与生物碎屑泥晶化有关,其有机质也来自微生物岩原生有机质。III和V型球粒的有机质成岩烘烤温度高,部分有机质可能来源于长期滞留在海水中的陆源有机物。其中,III型球粒形成可能与新生变形作用相关,V型则是生物碎屑泥晶化的产物。IV型球粒的有机质成岩烘烤温度中等,是微生物泥晶化和硅质碎屑混合的产物。 结论 二叠纪末大灭绝后,海洋中微生物爆发合成有机质,但只有少量成为颗粒有机碳沉降至海底并埋藏。微生物岩内部大多数泥晶球粒有机质均来源于微生物的躯体分解或代谢产物;而少量球粒的有机质来源于长期滞留在海水中的陆源有机质,它们经历多次成岩烘烤作用,具有较高的温度。成岩过程中有机质的烘烤及矿物重结晶作用对有机质的保存效率影响大,最终导致微生物岩中总有机碳含量较低。Abstract: Objective Following the mass extinction at the end of the Permian, microbialites were widely distributed in southern China, indicating a microbial bloom in the Lower Triassic following the cataclysm. However, microbialites generally exhibit low paleo-productivity, which seems to contradict the microbial explosion. This study analyzed the microbialites at the base of the Triassic Tianwan section in the Luodian area of Guizhou province, focusing on the petrological characteristics of micritic pellets and their thermal metamorphic evolution and sources of organic matter. The study clarifies the respective influences of microbially derived and seawater-trapped organic matter on pellet formation, with the aim of providing empirical evidence for exploring carbon cycling mechanisms in post-extinction microbialite systems. Methods In situ micro-area analytical techniques (optical microscopy, fluorescence microscopy, scanning electron microscopy (SEM) with energy-dispersive spectroscopy (EDS), and laser Raman spectroscopy) were used to conduct a detailed analysis of various types of peloids within the microbialites from the Tianwan section. These reveal the distribution patterns and thermal metamorphic evolutionary characteristics of internal organic matter. Results Based on morphology and infill materials, the various peloids were classified into six types: type I (microbially-induced peloid, MIP); type II (recrystallized microbially-induced peloid, RMIP); type III (aggregated detrital peloid, ADP); type IV (intraskeletal peloid, ISP); type V (dolomitized peloid, DP); and type VI (aggregated microbially-induced peloid, AMIP). Raman spectroscopy-derived metamorphic roasting temperatures provide insights into the thermal history and transformation of these peloids. The organic matter in types I, II, IV and VI (MIP, RMIP, ISP and AMIP) shows thermal maturity similar to that found in primary cyanobacteria in microbialites and shell fragments, indicating a common diagenetic history and microbial origin. Of these, peloids I, II and VI exhibit low thermal maturation temperatures, with organic matter mainly derived from the photosynthetic and metabolic processes of primary cyanobacteria and other microorganisms within microbialite systems. Specifically, peloid VI formed when micritic particles wrapped around peloid I, linked to bioclastic micritization. peloid IV shows moderate thermal maturation temperatures, resulting from a mix of microbial micritization and siliceous clastic material. By contrast, peloids III and V (ADP and DP) evidenced significantly higher metamorphic temperatures, suggesting multiple thermal alteration events. Peloid III may have been formed by neomorphism; peloid V was primarily shaped by bioclastic micritization. Conclusions In summary, after the end-Permian mass extinction, microbial blooms, especially cyanobacterial photosynthesis, led to the generation of large amounts of dissolved organic carbon (DOC) in the oceans. Due to frequent fluctuations in seawater redox conditions and rapid temperature increases, the DOC was typically oxidized into CO2 and released into the atmosphere, with only a small fraction being preserved as particulate organic carbon (POC) and deposited on the seafloor. Raman geothermometric analysis shows that most of the organic matter in the microbialite peloids, particularly in those associated with bioclastic peloidal envelopes, originated from the metabolic activity of primary cyanobacteria, which was the original organic matter in the microbialite system. A small portion of the organic matter may have been sourced from long-stored terrestrial organic matter in seawater, which underwent multiple diagenetic thermal alterations and shows higher thermal maturation temperatures. The thermal maturation of organic matter and mineral recrystallization during diagenesis had a significant impact on the efficiency of organic matter preservation, ultimately resulting in a low total organic carbon (TOC) content in the microbialites.
-
Key words:
- Permian-Triassic /
- South China /
- microbialite /
- peloid /
- organic origin
-
图 1 华南早三叠纪古地理图,显示田湾剖面的古地理背景(底图据冯增昭等,1997修改)
Figure 1. Early Triassic paleogeographical map of South China showing the setting of the Tianwan section (base map modified from Feng et al., 1997)
图 2 田湾剖面微生物岩序列的岩性柱状图以及典型岩性类型的野外露头
(a) Tianwan section, mainly showing the microbialite; (b) P-Tr boundary beds at the Tianwan section; (c) thrombolites within the microbialite of the Daye Formation; (d) coral-bearing bioclastic limestone of the Wujiaping Formation
Figure 2. Lithostratigraphy of the Tianwan section showing microbialite succession, and field outcrop photographs
Fig.2
图 3 田湾剖面微生物岩中观及微观特征
(a) polished surface of thrombolite; (b) polished surface of shell layers within microbialite; (c) polished surface of thrombolite with stylolite structures at microbialite base; (d⁃f) photomicrographs of thrombolite, displaying clotted textures; (g⁃i) photomicrographs of biological shell layers showing fossil fragments of ostracods, gastropods, bivalves and other organisms; white arrows indicate organic-matter-rich peloids; red arrows indicate clotted textures; yellow arrows indicate fossil fragments
Figure 3. Meso⁃microstructures of microbialites in the Tianwan section
Fig.3
图 4 田湾剖面微生物岩中微生物和后生生物化石的显微特征
(a-e) cyanobacteria Gakhumella, located within the sparry areas of thrombolite, where the matrix consists of sparry high-Mg calcite or dolomite; (a) clustered aggregates of Gakhumella with visible cup-like lamellae; (b) individual Gakhumella without spherical top; (c) top view of Gakhumella, showing the spherical calcite top and two rows of cup-like lamellae behind it; (d) aggregated area of Gakhumella, where some fossils display a double-sphere aggregation, representing cell division; Some fossils are well-preserved, showing the top and lamellae; (e) individual Gakhumella, with visible cup-like lamellae; (f) microconchids, overall tubular in appearance, and symbiotic with ostracods commonly found in the microbial carbonate shell layer; (g) gastropod fossils in microbial carbonate shell layer
Figure 4. Microscopic features of microbial and metazoan fossils within microbialites at the Tianwan section
Fig.4
图 5 I~III型球粒的光学显微镜下特征
(a) peloids I and II are partially retained in the same layer; some peloid II are deposited in the higher beds; (b, c) peloid I is black, mainly spherical in shape, with a particle size range 14-90 µm; It has indistinct boundaries and is distributed within calcite spar, containing abundant organic matter residues; The interior shows almost no recrystallized carbonate minerals; (d, e) peloid II is black and spherical, with a particle size range 10-130 µm; It has clear boundaries and an interior almost filled with recrystallized carbonate minerals, with a small amount of flocculent organic matter residue; (f-j) peloid III is circular or elliptical in shape; It is larger, with a particle size range 50-350 µm, with indistinct boundaries, and is mainly distributed in microbial micrite or calcite crystals; The particles are well-sorted and consist of fine-grained carbonate material
Figure 5. Photomicrographs of peloids I⁃III
Fig.5
图 6 IV~VI型球粒的光学显微镜下特征
(a⁃c) peloid IV has a particle size range 70-210 µm, is circular or elliptical in shape, with unclear boundaries; It is distributed within the cavities of bivalve fossils or between fossil fragments and is composed of micritic carbonate material; (d-f) peloid V is brown and angular in shape, with the largest particle size range 280-600 µm; It has clear boundaries, is often attached near fossil fragments, and some particles form larger aggregates; (g-j) peloid VI with a particle size range 70-300 µm has indistinct boundaries and is distributed around fossils within shell layers, with partial recrystallization
Figure 6. Photomicrographs of peloids IV⁃VI
Fig.6
图 7 不同类型球粒结构的粒径统计结果对比
The lower and upper bounds of each box represents the 25th percentile (Q1) and 75th percentile (Q3), respectively, The horizontal line inside the box indicates the median (Q2), The whiskers extend to the range of Q1-1.5×IQR (interquartile range) and Q3 + 1.5×IQR, The circle indicates the mean; Actual particle sizes are shown as scatter points to the right of each box
Figure 7. Comparison of particle size statistics of various peloids
Fig.7
图 8 不同激光照射下V和VI型球粒的荧光特征
(a) photomicrograph of peloid V in plane-polarized light (PPL); (b-e) fluorescence photographs of (a) in (b) green, (c) purple, (d) ultraviolet, and (e) blue light, to produce red, green, blue and yellow-green fluorescence, respectively; (f) photomicrograph of peloid VI in PPL; (g-j) fluorescence photographs of (f) in (g) green, (h) purple, (i) ultraviolet, and (j) blue light
Figure 8. Fluorescence photographs of peloids V and VI in different laser irradiation
Fig.8
图 9 不同球粒的能谱分析与元素组成对比
showing elemental mass percentage distributions, main elements Ca, O in left-hand scale; low-content elements in right-hand scale
Figure 9. EDS analysis and elemental composition comparisons of various peloids
Fig.9
图 10 不同类型有机质球粒结构的拉曼光谱点分析结果
(a-e) photomicrographs of a peloid under plane-polarized light; (f) Raman spectral point analysis for different types of peloids: spectral curves of different colors represent the Raman spectral results of the cross-filament position of the corresponding color
Figure 10. Raman spectral point analysis results for different types of organic matter spheroid structures
Fig.10
图 11 不同类型有机质球粒结构的拉曼光谱面分析结果
(a-c) photomicrographs of peloids I-III in PPL; (d-f) Raman spectral organic peak (1 600 cm-1) and calcite peak (1 080 cm-1); Raman spectral mappings of peloids I-III; (g⁃i) photomicrographs of peloids IV⁃VI in PPL; (j-l) Raman spectral organic peak (1 600 cm-1) and calcite peak (1 080 cm-1); Raman spectral mapping of peloids IV-VI
Figure 11. Raman spectral mapping of various peloids
Fig.11
图 12 研究剖面的球粒、生物化石及介壳碎片外部泥晶套的拉曼温度平面分布图
Results of calculations using formula 1; (a) peloid I, peak diagenetic temperature range 240 ℃-300 ℃; (b) peloid II, peak diagenetic temperature range 270 ℃-310 ℃; (c) peloid VI, peak diagenetic temperature range 170 ℃-310 ℃, with a low-temperature zone of 180 ℃-248 ℃ in the nucleus; (d) peloid III, peak diagenetic temperature range 260 ℃-340 ℃; (d) peloid V spherule, peak diagenetic temperature range 320 ℃-360 ℃; (f) peloid IV, peak diagenetic temperature range 220 ℃-350 ℃; (g) Gakhumella, peak diagenetic temperature range 300 ℃-360 ℃; (h) organic-rich micritic envelope, peak diagenetic temperature range 250 ℃-320 ℃; (i) foraminiferal fossils, peak diagenetic temperature range 270 ℃-360 ℃
Figure 12. Raman temperature maps for various peloids, fossils and micritic envelopes on the outside of shell fragments from the study section
Fig.12
图 13 不同类型球粒、生物化石和介壳碎片外部泥晶套的有机质石墨化程度分析
The background colors in the subfigures indicate different degrees of graphitization: pink. disordered organic matter; blue. early graphitization; light gray. intermediate graphitization; dark gray. advanced graphitization; In the scatter plot, darker colors represent R2 values close to 1, indicating better fitting results. Using the calculation based on formula (1). (1) as an example, most organic matter in the analyzed samples falls within the early graphitization stage: (a) peloid I (MIP) has most organic matter at the early stage of graphitization, with only a minor portion remaining in a disordered state; (b) peloid II (RMIP) shows poorly preserved, more scattered organic matter; (c) peloid III (ADP) contains some intermediate graphitization stage organic matter, with narrower Raman peak widths indicating a more ordered structure and higher thermal maturity; (d) peloid IV (ISP) with moderate graphitization, falling between different types; (e) peloid V (DP) with relatively dispersed distribution in temperature and peak width, but overall organic matter exhibits higher structural order and thermal maturity; (f) peloid VI (AMIP) has temperature and peak width ranges that almost overlap with types I and II, suggesting similar graphitization; (g) coccoid cyanobacterium Gakhumella has well-preserved organic matter along the laminar edges; (h) organic-rich micritic envelope shows significant organic matter enrichment; (i) foraminiferal fossil with relatively high organic matter structure
Figure 13. Graphitization degrees of organic matter preserved in various peloids, fossils and envelopes at the outside of the shell fragments
Fig.13 表 1 有机质拉曼温度计算公式
Table 1. Calformula for organic Raman temperature
公式编号 公式 适用温度范围/℃ 公式来源 描述 1 T=-2.15(FWHM.D1)+478 150~400 Kouketsu et al.,2014 基于D1峰的半峰宽(FWHM.D1)计算的温度 2 T=-6.78(FWHM.D2)+535 150~400 Kouketsu et al.,2014 基于D2峰的半峰宽(FWHM.D1)计算的温度 3 T=-445×R2+641 330~650 Beyssac et al.,2002 使用面积比参数R2[D1/(D1+D2+G)]计算温度 注: 公式1误差为±30 ℃,公式2与公式3误差为±50 ℃,单独使用某个公式受不同拉曼光谱参数对成岩烘烤温度的敏感性干扰,联合标定结果更加准确。 下载: 导出CSV
下载: 导出CSV
表 2 微生物岩中不同球粒的粒度、形态、矿物组成及光谱特征对比
Table 2. Comparisons of particle size, morphology, mineral composition and Raman spectral characteristics of various peloids within the microbialite
分类 I型球粒 II型球粒 III型球粒 IV型球粒 V型球粒 VI型球粒 特征 原生微生物诱导钙化球粒 重结晶微生物钙化球粒 陆源碎屑复合球粒 骨骼间填充球粒 白云岩化泥晶球粒 聚合型微生物诱导钙化球粒 粒度 细粒 细粒 中粒 中粒 粗粒 中粒 颗粒圆度 次圆状 次圆状 次棱角状 次圆状 棱角状 次棱角状、次圆状 分选性 良好 良好 一般 良好 一般 一般 基质矿物组成 亮晶方解石 亮晶方解石 泥晶方解石 泥晶方解石 亮晶白云石 亮晶白云石 胶结物类型 钙质泥晶胶结 钙质与少量白云质泥晶胶结 钙质泥晶胶结 钙质泥晶胶结 白云质泥晶胶结 白云质泥晶胶结 重结晶特征 无显著重结晶 核部重结晶显著,方解石为主 无显著重结晶 局部重结晶 局部重结晶 局部重结晶 分布位置 下部凝块石的方解石亮晶中 下部凝块石的方解石亮晶中 中下部凝块石的泥晶或亮晶区域 中部介壳层双壳化石内部/间隙 顶部介壳层生物化石碎片间隙 顶部介壳层生物化石腔体内部 颜色 黑色 黑色 棕色 黑色 深棕色 黑色 平均粒径/µm 45.3 50.5 151.5 125 291.4 175 粒径标准差/SD 15.6 7.3 53.6 31.6 114.3 52.1 粒径范围/µm 14~90 10~130 50~350 70~210 80~600 70~380 荧光特征 荧光响应弱 边缘荧光响应强 荧光响应弱 荧光响应弱 边缘荧光响应强,核部减弱 核部荧光响应强,边缘逐渐减弱 拉曼光谱特征 D1、G峰显著,可见D2、D4峰 D1、G峰清晰,可见方解石与白云石峰 D1、D2、G峰 D1、D2、G峰 D1、G峰显著,可见D2、D4峰,可见大量白云石峰 D1、D2、G峰显著,可见白云石峰 主要元素组成 富含Ca、O Ca和O含量最高,Mg含量略高 富含Ca、O,伴随Si、P、S少量富集 Ca和Si为主,少量Mg和Fe Mg/Ca显著升高,伴随Fe、S的富集 Mg/Ca显著升高,伴随P、Fe的富集
下载: 导出CSV
表 3 利用拉曼温度计对不同球粒、生物化石和介壳碎片泥晶套的有机质成岩烘烤温度计算结果
Table 3. Calculated peak period alternation temperatures of various peloids, fossils and micritic envelopes of shell fragments using Raman thermometer
类型 温度范围/℃ 集中分布范围/℃ 成岩烘烤程度 物质来源相似性 I型球粒 公式1:240~340公式2:310~380公式3:365~445 公式1:240~300公式2:310~330公式3:380~410 成岩烘烤程度最低 与VI型球粒和泥晶套接近 II型球粒 公式1:220~310公式2:210~430公式3:0~256 公式1:270~310公式2:210~320 公式3:64~256 成岩烘烤程度不均,核部重结晶严重,温度差异显著 与I型球粒有一定演化关系,但变质程度较高 III型球粒 公式1:260~340公式2:260~370公式3:330~500 公式1:260~340公式2:280~340公式3:400~460 高成岩烘烤程度,在公式2和公式3计算结果中尤为显著 与V型球粒物质来源最为接近 IV型球粒 公式1:220~350公式2:210~370公式3:350~470 公式1:220~280公式2:270~320公式3:390~430 公式1、公式2成岩烘烤程度较低,公式3中成岩烘烤程度中等 与Gakhumella和泥晶套接近 V型球粒 公式1:320~360公式2:250~380公式3:380~475 公式1:320~360公式2:286~350公式3:410~445 最高成岩烘烤程度,与III型球粒热演化规律一致 III型球粒和V型球粒的部分物质来源极为接近 VI型球粒 公式1:170~310公式2:250~370公式3:350~455 公式1:180~248公式2:260~330公式3:360~390 成岩烘烤程度最低,可能为I型球粒的演化产物 与I型球粒和泥晶套相似 Gakhumella 公式1:170~410公式2:255~340公式3:360~510 公式1:300~360公式2:280~330公式3:400~460 公式1和公式2成岩烘烤程度中等,公式3显示较高程度成岩烘烤 与后生生物化石接近 后生生物化石 公式1:220~385公式2:255~340公式3:330~465 公式1:270~360公式2:280~330公式3:425~455 公式1和公式2成岩烘烤程度中等,公式3显示较高程度成岩烘烤 与泥晶套成岩烘烤规律相近 泥晶套 公式1:250~320公式2:310~350公式3:380~435 公式1:250~320公式2:320~340公式3:400~425 低至中等成岩烘烤作用 与VI型球粒最为相似 注: 公式1与公式2计算结果相近,均表现为较低的成岩烘烤程度,与峰形拟合结果统一,公式3计算得到的温度范围与实际峰形所处温度范围差异较大,故主要参考公式1、2特征总结规律。
下载: 导出CSV
-
[1] 杜远生,黄虎,杨江海,等. 2013. 晚古生代—中三叠世右江盆地的格局和转换[J]. 地质论评,59(1):1-11. Du Yuansheng, Huang Hu, Yang Jianghai, et al. 2013. The basin translation from Late Paleozoic to Triassic of the Youjiang Basin and its tectonic signification[J]. Geological Review, 59(1): 1-11. [2] 冯增昭,鲍志东,吴胜和,等. 1997. 中国南方早中三叠世岩相古地理[J]. 地质科学,32(2):212-220. Feng Zengzhao, Bao Zhidong, Wu Shenghe, et al. 1997. Lithofacies palaeogeography of the Early and Middle Triassic of South China[J]. Chinese Journal of Geology, 32(2): 212-220. [3] 梁乐文,侯佳林,隋维康,等. 2023. 海洋沉积物微生物介导有机碳转化研究进展[J]. 微生物学报,63(5):1771-1786. Liang Lewen, Hou Jialin, Sui Weikang, et al. 2023. Research progress on microbiome and organic carbon transformation in marine sediment[J]. Acta Microbiologica Sinica, 63(5): 1771-1786. [4] 刘贝. 2023. 泥页岩中有机质:类型、热演化与有机孔隙[J]. 地球科学,48(12):4641-4657. Liu Bei. 2023. Organic matter in shales: Types, thermal evolution, and organic pores[J]. Earth Science, 48(12): 4641-4657. [5] 刘建波,江崎洋一,杨守仁,等. 2007. 贵州罗甸二叠纪末生物大灭绝事件后沉积的微生物岩的时代和沉积学特征[J]. 古地理学报,9(5):473-486. Liu Jianbo, Yoichi E, Yang Shouren, et al. 2007. Age and sedimentology of microbialites after the end-Permian mass extinction in Luodian, Guizhou province[J]. Journal of Palaeogeography, 9(5): 473-486. [6] 宋虎跃,童金南,田力,等. 2014. 南盘江地区二叠纪—三叠纪之交浅水台地古氧相研究[J]. 中国科学:地球科学,44(6):1273-1282. Song Huyue, Tong Jinnan, Tian Li, et al. 2014. Paleo-redox conditions across the Permian-Triassic boundary in shallow carbonate platform of the Nanpanjiang Basin, South China[J]. Science China Earth Sciences, 44(6): 1273-1282. [7] 田野,田云涛. 2020. 石墨化碳质物质拉曼光谱温度计原理与应用[J]. 地球科学进展,35(3):259-274. Tian Ye, Tian Yuntao. 2020. Fundamentals and applications of Raman spectroscopy of carbonaceous material (RSCM) thermometry[J]. Advances in Earth Science, 35(3): 259-274. [8] 王民, Li Zhongsheng. 2016. 激光拉曼技术评价沉积有机质热成熟度[J]. 石油学报,37(9):1129-1136. Wang Min, Li Zhongsheng. 2016. Thermal maturity evaluation of sedimentary organic matter using laser Raman spectroscopy[J]. Acta Petrolei Sinica, 37(9): 1129-1136. [9] 王永标,童金南,王家生,等. 2005. 华南二叠纪末大绝灭后的钙质微生物岩及古环境意义[J]. 科学通报,50(6):552-558. Wang Yongbiao, Tong Jinnan, Wang Jiasheng, et al. 2005. Calcareous microbialites and paleoenvironmental significance after the Late Permian mass extinction in South China[J]. Chinese Science Bulletin, 50(6): 552-558. [10] Adachi N, Asada Y, Ezaki Y, et al. 2017. Stromatolites near the Permian-Triassic boundary in Chongyang, Hubei province, South China: A geobiological window into palaeo-oceanic fluctuations following the end-Permian extinction[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 475: 55-69. [11] Adachi N, Ezaki Y, Liu J B. 2004. The fabrics and origins of peloids immediately after the end-Permian extinction, Guizhou province, South China[J]. Sedimentary Geology, 164(1/2): 161-178. [12] Alexandersson T. 1972. Intragranular growth of marine aragonite and Mg-calcite; evidence of precipitation from supersaturated seawater[J]. Journal of Sedimentary Research, 42(2): 441-460. [13] Algeo T J, Henderson C M, Tong J N, et al. 2013. Plankton and productivity during the Permian-Triassic boundary crisis: An analysis of organic carbon fluxes[J]. Global and Planetary Change, 105: 52-67. [14] Beyssac O, Goffé B, Chopin C, et al. 2002. Raman spectra of carbonaceous material in metasediments: A new geothermometer[J]. Journal of Metamorphic Geology, 20(9): 859-871. [15] Broughton P L. 2023. Morphogenesis of crystal fan fabrics of the Wolfenden cool water tufa deposit in western Canada[J]. Facies, 69(1): 3. [16] Canfield D E, van Zuilen M A, Nabhan S, et al. 2021. Petrographic carbon in ancient sediments constrains Proterozoic Era atmospheric oxygen levels[J]. Proceedings of the National Academy of Sciences of the United States of America, 118(23): e2101544118. [17] Carlson C A, Hansell D A. 2015. sources DOM, sinks, reactivity, and budgets[M]//Hansell D A, Carlson C A. Biogeochemistry of marine dissolved organic matter. Amsterdam: Elsevier: 65-126. [18] Chafetz H S. 1986. Marine peloids; a product of bacterially induced precipitation of calcite[J]. Journal of Sedimentary Research, 56(6): 812-817. [19] Chen Z Q, Benton M J. 2012. The timing and pattern of biotic recovery following the end-Permian mass extinction[J]. Nature Geoscience, 5(6): 375-383. [20] Chen Z Q, Fang Y H, Wignall P B, et al. 2022. Microbial blooms triggered pyrite framboid enrichment and oxygen depletion in carbonate platforms immediately after the latest Permian extinction[J]. Geophysical Research Letters, 49(7): e2021GL096998. [21] Chen Z Q, Tu C Y, Pei Y, et al. 2019. Biosedimentological features of major microbe-metazoan transitions (MMTs) from Precambrian to Cenozoic[J]. Earth-Science Reviews, 189: 21-50. [22] Derrien M, Jeanneau L, Jardé E, et al. 2023. Exploration of changes in the chemical composition of sedimentary organic matter and the underlying processes during biodegradation through advanced analytical techniques[J]. Environmental Chemistry, 20(5): 212-225. [23] Dufresne W J B, Rufledt C J, Marshall C P. 2018. Raman spectroscopy of the eight natural carbonate minerals of calcite structure[J]. Journal of Raman Spectroscopy, 49(12): 1999-2007. [24] Erwin D H. 1994. The Permo-Triassic extinction[J]. Nature, 367: 231-236. [25] Ezaki Y, Liu J B, Nagano T, et al. 2008. Geobiological aspects of the earliest Triassic microbialites along the southern periphery of the tropical Yangtze Platform: Initiation and cessation of a microbial regime[J]. Palaios, 23(6): 356-369. [26] Fang Y H, Chen Z Q, Kershaw S, et al. 2017. Permian-Triassic boundary microbialites at Zuodeng section, Guangxi province, South China: Geobiology and palaeoceanographic implications[J]. Global and Planetary Change, 152: 115-128. [27] Flügel E. 2004. Microfacies of carbonate rocks: Analysis, interpretation and application[M]. Heidelberg, Berlin/New York: Springer: 1-976. [28] Foster W J, Lehrmann D J, Yu M, et al. 2018. Persistent environmental stress delayed the recovery of marine communities in the aftermath of the latest Permian mass extinction[J]. Paleoceanography and Paleoclimatology, 33(4): 338-353. [29] Gale L, Skaberne D, Peybernes C, et al. 2016. Carnian reefal blocks in the Slovenian Basin, eastern southern Alps[J]. Facies, 62(4): 23. [30] Heindel K, Foster W J, Richoz S, et al. 2018. The formation of microbial-metazoan bioherms and biostromes following the latest Permian mass extinction[J]. Gondwana Research, 61: 187-202. [31] Hong H L, Fang Q, Zhao L L, et al. 2017. Weathering and alteration of volcanic ashes in various depositional settings during the Permian-Triassic transition in South China: Mineralogical, elemental and isotopic approaches[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 486: 46-57. [32] Kershaw S, Crasquin S, Li Y, et al. 2012. Microbialites and global environmental change across the Permian-Triassic boundary: A synthesis[J]. Geobiology, 10(1): 25-47. [33] Kouketsu Y, Mizukami T, Mori H, et al. 2014. A new approach to develop the Raman carbonaceous material geothermometer for low‐grade metamorphism using peak width[J]. Island Arc, 23(1): 33-50. [34] Luo G M, Wang Y B, Grice K, et al. 2013. Microbial-algal community changes during the latest Permian ecological crisis: Evidence from lipid biomarkers at Cili, South China[J]. Global and Planetary Change, 105: 36-51. [35] Macintyre I G. 1985. Submarine cements-the peloidal question[M]//Schneidermann N, Harris P M. Carbonate cements: Based on a symposium sponsored by the society of economic paleontologists and mineralogists. Tulsa: SEPM Society for Sedimentary Geology: 508-508. [36] Megonigal J P, Hines M E, Visscher P T. 2004. Anaerobic metabolism: Linkages to trace gases and aerobic processes[M]//Schlesinger W H. Biogeochemistry. Oxford: Elsevier-Pergamon: 1-18. [37] Monty C L V. 1976. The origin and development of cryptalgal fabrics[J]. Developments in sedimentology, 20: 193-249. [38] Neuweiler F. 1993. Development of Albian microbialites and micro-bialite reefs at marginal platform areas of the Vasco-Cantabrian Basin (Soba reef area, Cantabria, N. Spain)[J]. Facies, 29(1): 231-249. [39] Pei Y, Chen Z Q, Fang Y H, et al. 2019. Volcanism, redox conditions, and microbialite growth linked with the end-Permian mass extinction: Evidence from the Xiajiacao section (western Hubei province), South China[J]. Palaeogeography, Palaeoclimatology, Palaeo-ecology, 519: 194-208. [40] Pei Y, Duda J P, Reitner J. 2021. Sedimentary factories and ecosystem change across the Permian-Triassic Critical Interval (P-TrCI): Insights from the Xiakou area (South China)[J]. PalZ, 95(4): 709-725. [41] Philp R P. 1985. Petroleum formation and occurrence[J]. Eos, Transactions American Geophysical Union, 66(37): 643-644. [42] Reid R P. 1987. Nonskeletal peloidal precipitates in Upper Triassic reefs, Yukon Territory (Canada)[J]. Journal of Sedimentary Research, 57(5): 893-900. [43] Samankassou E, Tresch J, Strasser A. 2005. Origin of peloids in Early Cretaceous deposits, Dorset, South England[J]. Facies, 51(1/2/3/4): 264-274. [44] Sepkoski J J. 1984. A kinetic model of Phanerozoic taxonomic diversity. III. Post-Paleozoic families and mass extinctions[J]. Paleo-biology, 10(2): 246-267. [45] Shen J, Schoepfer S D, Feng Q L, et al. 2015. Marine productivity changes during the end-Permian crisis and Early Triassic recovery[J]. Earth-Science Reviews, 149: 136-162. [46] Sparkes R, Hovius N, Galy A, et al. 2013. Automated analysis of carbon in powdered geological and environmental samples by Raman spectroscopy[J]. Applied Spectroscopy, 67(7): 779-788. [47] Sparkes R B, Hovius N, Galy A, et al. 2020. Survival of graphitized petrogenic organic carbon through multiple erosional cycles[J]. Earth and Planetary Science Letters, 531: 115992. [48] Steinhauff D M, Abubshait A, Purkis S J. 2021. Red Sea Holocene carbonates: Windward platform margin and lagoon near Al-Wajh, northern Saudi Arabia[J]. Journal of Sedimentary Research, 91(8): 847-875. [49] Sun S Q, Wright V P. 1989. Peloidal fabrics in Upper Jurassic reefal limestones, Weald Basin, southern England[J]. Sedimentary Ge-ology, 65(1/2): 165-181. [50] Sun Y D, Joachimski M M, Wignall P B, et al. 2012. Lethally hot temperatures during the Early Triassic greenhouse[J]. Science, 338(6105): 366-370. [51] Tang H, Kershaw S, Liu H, et al. 2017. Permian-Triassic boundary microbialites (PTBMs) in Southwest China: Implications for paleoenvironment reconstruction[J]. Facies, 63(1): 2. [52] Wada S, Aoki M N, Tsuchiya Y, et al. 2007. Quantitative and qualitative analyses of dissolved organic matter released from Ecklonia cava Kjellman, in Oura Bay, Shimoda, Izu Peninsula, Japan[J]. Journal of Experimental Marine Biology and Ecology, 349(2): 344-358. [53] Wu S Q, Chen Z Q, Fang Y H, et al. 2017. A Permian-Triassic boundary microbialite deposit from the eastern Yangtze Platform (Jiangxi province, South China): Geobiologic features, ecosystem composition and redox conditions[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 486: 58-73. [54] Wu S Q, Chen Z Q, Fang Y H, et al. 2022. Benthic Pleurocapsales (Cyanobacteria) blooms catalyzing carbonate precipitation and dolomitization following the end-Permian mass extinction[J]. Geophysical Research Letters, 49(24): e2022GL100819. [55] Wu S Q, Reitner J, Harper D A T, et al. 2024. New keratose sponges after the end-Permian extinction provide insights into biotic recoveries[J]. Geobiology, 22(1): e12582. [56] Yuan D X, Zhang Y C, Shen S Z. 2018. Conodont succession and reassessment of major events around the Permian-Triassic boundary at the Selong Xishan section, southern Tibet, China[J]. Global and Planetary Change, 161: 194-210. [57] Zheng Z J, Chen Z Q, Grasby S E, et al. 2024. Carbon-sulfur isotope and major and trace element variations across the Permian-Triassic boundary on a shallow platform setting (Xiejiacao, South China)[J]. Chemical Geology, 657: 122115. [58] Zhou K, Pratt B R. 2019. Upper Devonian (Frasnian) stromatactis-bearing mud mounds, western Alberta, Canada: Reef framework dominated by peloidal microcrystalline calcite[J]. Journal of Sedimentary Research, 89(9): 833-848. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 247
- HTML全文浏览量: 49
- PDF下载量: 174
- 被引次数: 0

