

 下载:
下载:







-
遗迹化石是造迹生物与沉积底质相互作用的产物,其造迹生物的非限定性、原地保存性、量大且易于识别等特征,使得遗迹化石成为探索古生物和古环境相互作用的有力工具(Knaust and Bromley,2012;齐永安等,2012;罗茂等,2021;许晴旸等,2023)。特别是穴居生物对其栖息地环境变化反应敏感,因此,在很多情况下,遗迹化石比实体化石更能反映沉积环境条件(Wetzel and Rodríguez-Tovar,2021)。遗迹化石的种属、丰度、个体大小、穿透沉积物的深度、潜穴壁的光滑度和结构等,均具有较高的环境指示意义(Wetzel and Rodríguez-Tovar,2021)。例如,个体细小的、浅层分布的规则图案状的潜穴系统通常反映安静缺氧的深水环境(Uchman and Wetzel,2012),而个体较大、形态简单、丰度高的垂直潜穴一般代表动荡富氧的浅水环境(Buatois and Mángano,2004,2011);潜穴壁光滑通常指示沉积底质具有一定的固结度(Knaust,2021)。遗迹化石Thalassinoides Ehrenberg(1944)是最古老的复杂遗迹化石,地史分布从寒武纪至今(Jin et al.,2012,2013;Knaust,2021),但主要集中在中生代和新生代(Bromley and Ekdale,1984;Carmona et al.,2004;El-Sabbagh et al.,2017)。其典型特征是具有大量T-形或Y-形分支的网状或三维箱状开放潜穴系统。Thalassinoides在多种岩性中均可发育,包括灰岩、白云岩、砂岩、粉砂岩和泥岩(Sheehan and Schiefelbein,1984;Joseph et al.,2012;Sharafi et al.,2012;Leonowicz,2015;Marusin et al.,2022),广泛应用在沉积环境解释(Ekdale and Bromley,2003;李凯楠等,2019;Knaust,2024;Zhang et al.,2024)和海洋底栖生态系统工程研究中(McIlroy and Logan,1999;Solan and Kennedy,2002;Erwin,2008;Liu et al.,2024)。
河南寒武系发育大量的Thalassinoides,地层跨度从寒武系黔东统都匀阶朱砂洞组到芙蓉统排碧阶崮山组,形态特征从二维网状到三维箱状展布,同时伴有潜穴结构、直径、大小、扰动强度、扰动深度等的变化。本文通过对河南富含Thalassinoides的地层沉积学特征及对应的不同遗迹种的特征研究,探索Thalassinoides不同遗迹种与沉积环境之间的关系,为更好地利用Thalassinoides解释古环境及重建寒武纪古生态提供支撑材料。
-
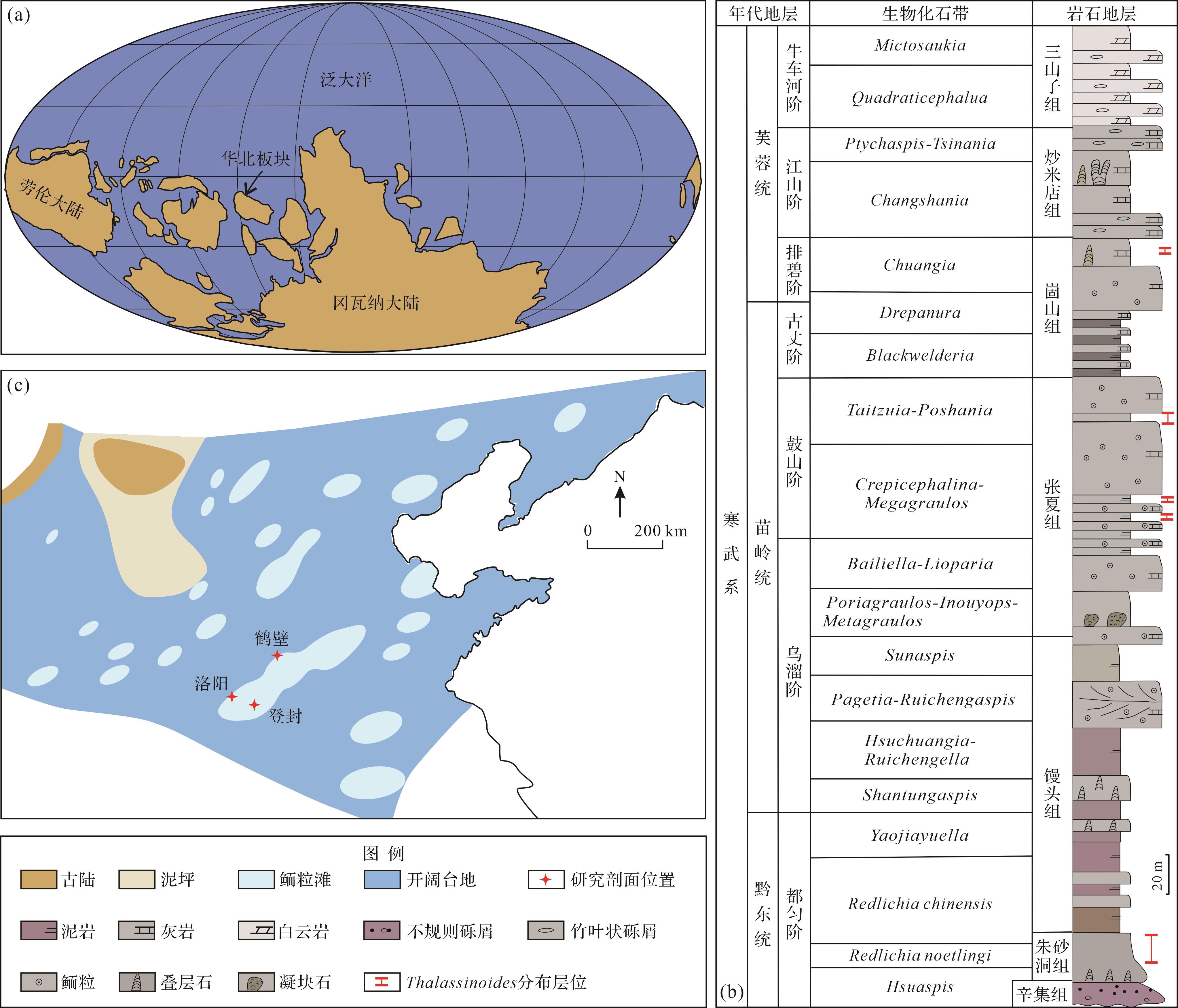
寒武纪华北板块位于赤道附近,是一个被泛海洋包围的孤立克拉通(图1a,Torsvik and Cocks,2019)。寒武纪之初华北板块为古陆,长期遭受风化剥蚀,黔东世都匀期被水淹没开始接受沉积(冯增昭等,1989,2004;裴放等,2008),整体表现为西高东低、南高北低的古地理格局,且以开阔台地相、浅滩和潮坪沉积为主(牟传龙等,2016)。华北板块南部河南省寒武系除了缺失底部纽芬兰统和黔东统筇竹寺阶外,其余地层出露良好且连续,自下而上沉积了黔东统辛集组、朱砂洞组和馒头组下段,苗岭统馒头组上段、张夏组和崮山组下段,以及芙蓉统崮山组上段、炒米店组和三山子组(图1b)。本文遗迹化石Thalassinoides的主要剖面位置分布在河南省登封朱砂洞组、洛阳张夏组和鹤壁崮山组(图1c)。

图 1 (a)寒武纪苗岭世华北板块的古地理位置(据Torsvik and Cocks,2019修改);(b)河南寒武纪的综合地层图(据裴放,2000;朱茂炎等,2019修改);(c)寒武纪苗岭世华北板块的古地理格局(据冯增昭等,2002修改)
Figure 1. (a) Paleogeographic location of the North China Block in the Cambrian Miaolingian (modified from Torsvik and Cocks, 2019); (b) synthetical strata of the Cambrian in Henan (modified from Pei, 2000; Zhu et al., 2019); (c) paleogeographic pattern in the CambrianMiaolingian of the North China Block (modified from Feng et al., 2002)
-
对发育Thalassinoides的地层进行详细的观察与测量,识别出重要的沉积学和遗迹学特征。沉积学特征主要包括岩性和岩性组合、沉积构造、沉积微相和伴生的生物成因沉积构造等。遗迹学特征主要包括遗迹化石的形态、产状、直径、扰动深度、填充物、衬壁的发育情况、生物扰动强度等。其中,生物扰动强度的划分依据Droser and Bottjer(1986)提出的生物扰动指数(BI):生物扰动构造对原始沉积组构的改造程度的比例,划分为6个等级,分别对应不同的扰动量:1(0%),2(1%~10%),3(10%~40%),4(40%~60%),5(>60%),6(100%)。
-
登封朱砂洞组厚35.2 m,根据岩性特征分为三段(图2):底部一段厚7.2 m,以浅黄灰色膏溶角砾岩开始,角砾的大小不等,成分为具水平纹层的灰岩(图2a);向上过渡到浅灰色厚层具水平纹层的灰质白云岩(图2b,c),夹一层侧向不连续的叠层石丘(图2b底部);最上部为一层发育小型低角度交错层理的细颗粒灰岩(图2e),显微镜下显示颗粒具有一定的分选性和磨圆度(图2f)。中部二段厚14.5 m,主要为深灰色厚层豹斑灰岩(图3),发育大量的遗迹化石Thalassinoides suevicus。风化面上豹斑呈深灰色或土黄色,凸出于层面之上;基质呈灰白色;使用浓度5%的稀盐酸滴在风化面上,豹斑起泡微弱,而基质部分起泡剧烈;中间夹5层竹叶状砾屑灰岩,砾屑呈扁平状,定向性不明显(图2d)。顶部三段厚13.5 m,主要为浅灰色藻纹层灰质白云岩(图2g,h)夹浅紫红色灰质泥岩,白云岩中发育鸟眼构造。

图 2 登封地区朱砂洞组综合柱状图及野外和显微镜照片
Figure 2. Synthetical strata, field and microscopic photos of Zhushadong Formation in Dengfeng area

图 3 登封地区朱砂洞组Thalassinoides suevicus野外和显微镜照片
Figure 3. Field and microscopic photos of Thalassinoide suevicus from the Zhushadong Formation in the Dengfeng area
-
二段的豹斑构造主要起因于生物扰动构造Thalassinoides suevicus。T. suevicus为在层面上多分支的潜穴系统(图3a,b),分支角度以Y-形分支为主。潜穴直径通常为3~8 mm,且在分支点处呈现明显的膨大,膨大处一般宽10~13 mm。T. suevicus常密集分布,布满整个层面,潜穴之间的交叉和叠覆现象常见。剖面上呈现出不规则的斑状构造,偶见分支现象(图3d),扰动深度较浅,一般不超过3 cm(图3c,d)。潜穴的填充物通常呈现出灰色(图3a)、土黄色(图3b,d)和深灰色(图3c),与浅灰色的围岩形成较为鲜明的对比。显微镜下观察潜穴填充物以细晶白云石为主,而围岩为泥晶方解石,且潜穴与围岩的边界清晰(图3e,f)。生物扰动指数BI为2~5(图3a~d),常见3~4,在二段的中部最强(图2)。
-
洛阳张夏组厚133 m,根据岩性特征分为四段(图4)。底部一段厚15.8 m,主要由浅灰色薄板状灰岩与极薄层状泥岩组成(图4a),中间夹四层竹叶状砾屑灰岩(图4a)。下部二段厚20.2 m,主要为微生物岩和生物扰动灰岩(图4b,c)及一层夹有薄层泥晶灰岩的鲕粒灰岩,生物扰动构造为Balanoglossites triadicus。微生物岩主要为叠层石(图4b)和凝块石(图4c,d)。中部三段厚35 m,为颗粒灰岩和生物扰动灰岩互层(图4e,f),生物扰动构造为Thalassinoides horizontalis(图5)。上部四段厚62 m,以厚层状鲕粒灰岩为主(图4g),夹数层侧向不连续的生物扰动灰岩(图4g);鲕粒灰岩的鲕粒大小均匀,多呈放射状纹层结构,颗粒接触,胶结物为亮晶方解石(图4h)。

图 4 洛阳地区张夏组综合柱状图及野外和显微镜照片
Figure 4. Synthetical strata, field and microscopic photos of Zhangxia Formation in Luoyang area

图 5 洛阳地区张夏组Thalassinoides野外和显微镜照片
Figure 5. Field and microscopic photos of Thalassinoides from the Zhangxia Formation in the Luoyang area
-
张夏组发育大量的遗迹化石,其中Thalassinoides horizontalis分布在三段下部,Thalassinoides bacae分布在四段上部。Thalassinoides horizontalis 的形态特征表现为层面多分支的二维潜穴。根据分支形态、分支间距和潜穴直径的不同,分为两个类型。T. horizontalis Type 1(图5a),为Y-形分支,分支间距较长,2~30 cm,多为15~30 cm,直径8~12 mm,层面上呈稀疏的管状结构,没有交叉和叠覆,扰动强度低,一般为2;分布在薄板状灰岩的层面上,不同层之间的潜穴个体和形态差别不大(图5b)。与T. horizontalis Type 1伴生的有大量的生物碎屑,碎屑个体较大,主要为三叶虫碎片。T. horizontalis Type 2分布在T. horizontalis Type 1之上的地层和较厚层灰岩层面上,呈T-形分支,分支间距小,单支长度2~4 cm,潜穴直径为7~8 mm,层面上呈现网状结构(图5c),生物扰动强度高,BI为3,且潜穴管内常伴生有铁矿化作用(图5d);围岩为较细的泥晶方解石(图5d)。Thalassinoides bacae分布在四段顶部鲕粒灰岩之间的薄层泥晶灰岩中,呈现多分支、具多个垂直管道的箱状潜穴系统(图5e~g),水平截面上多成圆点状或长条带状,偶有分支(图5g),剖面上呈倾斜的或垂直的管状、椭圆状等(图5e,f),扰动深度较深,约为15 cm;潜穴具有较厚的衬壁,衬壁为极细的泥晶方解石,衬壁外面通常围绕着白云石晕圈,而围岩由较细的泥晶方解石组成(图5h)。
-
鹤壁崮山组厚21 m,根据岩性特征分为三段(图6),下部一段厚12.1 m,主要为浅灰色—深灰色薄层泥晶灰岩与灰黄色泥岩互层(图6a,e),夹数层竹叶状砾屑灰岩和薄层鲕粒灰岩(图6b),薄层泥晶灰岩中含有极其细小的遗迹化石Mycellia(图6c)和极其细小的生物碎屑(图6f),黄色泥岩层面上有三叶虫碎片(图6d)和黄铁矿晶体(图6f)。中部二段厚5.5 m,主要为竹叶状砾屑灰岩,鲕粒灰岩和浅灰色叠层石丘灰岩,叠层石丘横向上不连续,以大型柱状结构为主,具抗浪格架,柱间充填大颗粒碎屑,底部有厚度不均匀的砾屑层。上部三段厚3.4 m,为深灰色生物扰动泥晶灰岩夹极薄灰黄色泥线(图6e),发育多层砾屑灰岩和微生物丘灰岩(图6g,h)。

图 6 鹤壁地区崮山组综合柱状图及野外和显微镜照片
Figure 6. Synthetical strata, field and microscopic photos of the Gushan Formation in the Hebi area
-
鹤壁崮山组三段薄层泥晶灰岩中发育Thalassinoide bacae。T. bacae作为单一遗迹属种占领地层,在层面上呈串珠状(图7a)或稀疏的管状突起(图7b),以Y-形分支为主(图7c),含有少量T-形分支(图7c)。T. bacae发育厚衬壁,露头暴露在外长期遭受风化作用使得晕圈和填充物被完全剥蚀,剖面上衬壁更加突出(图7d)。潜穴直径为5~8 mm。T. bacae在层面和剖面上的扰动指数较低,BI为2~3,扰动深度一般小于5 cm。显微镜下观察到填充物和晕圈以细晶白云石为主,衬壁和围岩成分为微晶方解石(图7e,f)。

图 7 鹤壁地区崮山组Thalassinoides bacae野外和显微镜照片
Figure 7. Field and microscopic photos of Thalassinoidesbacae from the Gushan Formation in the Hebi area
-
同一遗迹属通常是同一类造迹生物或者营相同生态习性的生物所建造(Seilacher,2007),而遗迹种的不同,一般受底质性质、沉积速率、水动力条件、营养物质和氧含量等多方面因素影响(Taylor et al.,2003;Feng et al.,2019)。Thalassinoides作为浅海沉积中常见的遗迹化石,其遗迹种或形态的细微变化与沉积环境有密切的关系(Knaust,2024;Zhang et al.,2024)。河南寒武系数套地层中均发育Thalassinoides,共包括三个遗迹种的四个形态类型,将其与所在的沉积环境相结合,探究两者之间的关系。
登封朱砂洞组一段主体岩性为灰岩和灰质白云岩,发育水平纹层,底部的膏溶角砾岩是暴露在干旱的条件下又经大气降水淋滤而形成(彭勇民等,2012),低角度双向交错层理是潮汐作用的产物,表明该段形成在潮上带。二段为一套生物扰动灰岩,夹竹叶状砾屑灰岩,形成在完全水下的潮间带,时常受到风暴影响。Thalassinoides suevicus呈现浅阶层特征、密度极高、叠覆现象普遍,指示营养物质和含氧量充足的环境,大量的造迹生物在沉积物浅层挖掘潜穴、进食(Griffis and Suchanek,1991;Dworschak and Rodrigues,1997)。潜穴形态不规则,且潜穴边界与围岩较为清晰,结合竹叶状砾屑灰岩的出现,说明表层沉积物具有一定的固结性(Ekdale and Bromley,2003;Ekdale and Lamond,2003)。三段藻纹层灰质白云岩夹浅紫红色灰质泥岩,发育鸟眼构造,说明水体变浅,蒸发作用强,为潮上带。
华北板块在张夏期早期发生最大的海侵事件,然后进入强迫型海退阶段(冯增昭等,2004)。洛阳张夏组一段深灰色泥岩和页岩发育,缺乏生物扰动,偶尔有竹叶状砾屑层,表明沉积环境为潮下带深水区,水深在风暴浪基面附近,总体上水体能量较低,但偶尔受到风暴的影响。二段以富含生物碎屑的微生物岩和含有少量Balanoglossites triadicus的扰动灰岩为主,说明水体深度变浅,水体氧含量有所增加,为潮下带浅水区。三段下部是颗粒灰岩和富含Thalassinoides horizontalis Type 1的薄板状灰岩,形成环境为鲕粒滩的滩前浅水区,水动力条件强,常受波浪或湍流的影响,从而改善了水体循环,带来氧气和营养物质等,形成直径更大,潜穴管更长的洞穴系统,且沉积速率快,生物尚未对沉积物进行充分扰动。而Thalassinoides horizontalis Type 2保存在较厚层状微晶灰岩中,该层生物碎屑少,生物扰动强度大,但是潜穴个体较Thalassinoides horizontalis Type 1小,分支的距离小,潜穴壁更加光滑,说明沉积环境较稳定,殖居窗长(Ekdale and Bromley,2003)。沉积速率缓慢,生物可对沉积物进行长时间的扰动,且沉积底质具有一定的固结性(Scholle and Ulmer-Scholle,2003;Rodríguez-Tovar et al.,2017;Wang et al.,2019)。其形成环境位置较T. horizontalis Type 1略深,水体动能略低,氧含量降低。
洛阳张夏组顶部四段以厚层鲕粒灰岩为主,鲕粒的分选性好,鲕粒间为亮晶方解石胶结,说明形成在持续高能动荡的鲕粒滩环境。T. bacae发育在厚层鲕粒灰岩的微晶灰岩夹层中,且有些夹层横向上呈向下凹的透镜体状,说明其形成环境与鲕粒滩关系密切,可能为高能鲕粒滩的滩间局限海,属于半封闭环境,水体较安静且氧含量低。T. bacae垂向管道多且较深,向下延伸到一定深度后分支,形成复杂的三维箱状结构。潜穴有致密且较厚的衬壁体现了造迹生物具有极强的建造潜穴能力以及适应营养物质和氧气匮乏的能力(Ekdale and Bromley,2003;李凯楠等,2019)。
鹤壁崮山组一段主要为浅灰色—深灰色薄层泥晶灰岩与灰黄色泥岩互层,是潮下带深水环境的产物,而极其细小的遗迹化石Mycellia是典型低能缺氧环境中的代表,加之薄片中普遍存在黄铁矿晶体,也进一步证实了沉积环境的缺氧。二段竹叶状砾屑灰岩,鲕粒灰岩和叠层石丘灰岩,含生物扰动构造,说明二段的水体变浅,为潮下带浅水区。上部三段砾屑灰岩、微生物丘灰岩和含T. bacae的生物扰动灰岩,生物扰动强度较二段低,说明水体加深。尽管崮山组的T. bacae的形态特征与张夏组类似,但潜穴密度较张夏组低,可能由两者的沉积环境决定的,崮山组水深比张夏组上部深,含氧量较张夏组更低。
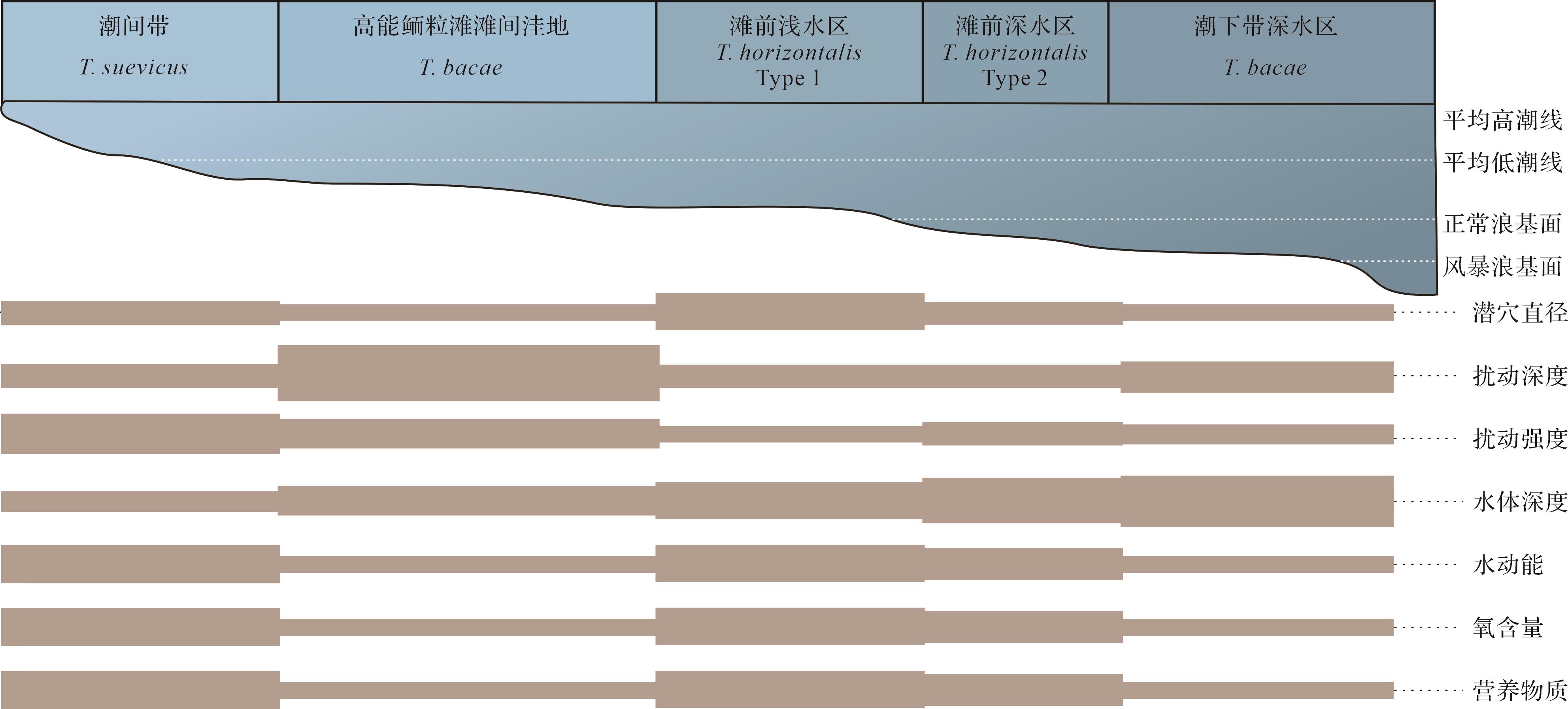
综上所述,河南寒武系Thalassinoides的形态变化与沉积环境有密切的关系,在氧气和营养物质充足的潮间带发育大量的T. suevicus,而在沉积速率快、氧气和营养物质充足的鲕粒滩滩前环境则发育个体大、分支长的浅层水平潜穴T. horizontalis Type 1;随着水体向海略微加深,过渡到密集分支、直径较小的T. horizontalis Type 2;在半封闭的缺少营养物质和氧气的静水环境中是机会种生物建造的发育厚壁的T. bacae(图8)。从寒武纪初期到寒武纪中后期,Thalassinoides的形态逐渐变得复杂,从浅阶层到深阶层、从近表层的二维潜穴逐渐演变为复杂的三维潜穴系统、从没有衬壁到发育致密的厚衬壁,反映了造迹生物应对不同生存环境采取的建筑策略以及行为习性的进化。

图 8 河南寒武系不同形态的Thalassinoides与沉积环境的关系模式图
Figure 8. Relationship schematic model between different forms of Thalassinoides and
various paleoenvironmental factors from the Cambrian in Henan province -
河南寒武系发育大量的Thalassinoides,共识别出了三个遗迹种的四个形态类型:登封朱砂洞组的T. suevicus,洛阳张夏组的T. bacae、T. horizontalis Type 1和Type 2,鹤壁崮山组的T. bacae。登封朱砂洞组T. suevicus是Y-形分支的(分支处膨大)浅阶层网状潜穴系统,密集分布,形成于水体动荡、氧气和营养物质充足的潮间带。洛阳张夏组三段T. horizontalis Type 1以Y-形分支、分支间距长、直径大为特征,稀疏分布,形成于水动力强,氧气和营养物质丰富、沉积速率高的鲕粒滩的滩前浅水区,T. horizontalis Type 2以T-形分支、分支间距短、直径略小,扰动强度大为特征,形成于鲕粒滩的滩前较深水区。洛阳张夏组四段和崮山组三段的T. bacae为具有垂向管并发育厚衬壁的三维箱状潜穴系统,扰动强度低到中等,形成于半闭塞的滩间洼地、滩间局限海或缺氧的深水潮下带。
Trace Fossil Thalassinoides and Its Paleoenvironmental Factors from the Cambrian in Henan Province
-
摘要: 目的 河南寒武系发育大量遗迹化石Thalassinoides,通过研究遗迹种的不同形态和分布特征,分析其沉积环境控制因素。 方法 基于沉积学和遗迹学研究,将该区Thalassinoides形态、大小、扰动深度、扰动强度等特征与沉积环境参数相结合。 结果 密集分布的Y-形分支、分支处膨大的二维潜穴Thalassinoides suevicus发育在水体动荡氧气充足的潮间带;稀疏分布的Y-形长分支、直径大的二维潜穴Thalassinoides horizontalis Type 1形成于水体动荡、氧气充足、营养物质丰富的鲕粒滩滩前浅水区;较密集分布的T-形分支、分支短直径小的二维潜穴Thalassinoides horizontalis Type 2形成在鲕粒滩滩前深水区;具有垂直管道、多分支且发育厚衬壁的三维箱状潜穴Thalassinoides bacae则发育在氧和营养物质均匮乏的滩间局限海和深水潮下带。 结论 Thalassinoides不同遗迹种及形态的变化受控于沉积环境的水深、水动能、营养物质和氧含量等因素,造迹生物采取不同的策略以应对变化的沉积环境。Abstract: Objective A large number of Thalassinoides trace fossils developed in the Cambrian strata of Henan province. We studied the morphology and distribution characteristics of different ichnospecies and analyzed the sedimentary environment-controlling factors. Methods Based on lithology and ichnography, the morphology, size, disturbance depth, and disturbance intensity of Thalassinoides in this area were combined with sedimentary environment parameters. Results The dense network Thalassinoides suevicus features
Y-shaped branches with enlarged intersections, developed in the intertidal zone where water is turbulent and oxygen is abundant. Sparse Thalassinoides horizontalis Type 1, which is Y-shaped long branches with large diameters, formed in the shallow water area in front of the oolitic beach with abundant oxygen and nutrients due to water turbulence. Thalassinoides horizontalis Type 2, T-shaped short branches with smaller diameters, occurred in deeper waters on the seaward side in front of the oolitic beach. The three-dimensional, boxwork burrows of Thalassinoides bacae, featuring vertical tubes, branches, and thick lining walls, developed in interbeach-confined seas and deep subtidal zones where oxygen and nutrients were deficient. Conclusions The variation in different ichnospecies and morphologies of Thalassinoides is controlled by factors such as water depth, water kinetic energy, nutrients, and oxygen content of the sedimentary environment. The trace maker adopted different strategies to cope with the changing sedimentary environment. -
Key words:
- Cambrian /
- carbonate platform /
- bioturbation /
- sedimentary environment
-
图 1 (a)寒武纪苗岭世华北板块的古地理位置(据Torsvik and Cocks,2019修改);(b)河南寒武纪的综合地层图(据裴放,2000;朱茂炎等,2019修改);(c)寒武纪苗岭世华北板块的古地理格局(据冯增昭等,2002修改)
Figure 1. (a) Paleogeographic location of the North China Block in the Cambrian Miaolingian (modified from Torsvik and Cocks, 2019); (b) synthetical strata of the Cambrian in Henan (modified from Pei, 2000; Zhu et al., 2019); (c) paleogeographic pattern in the CambrianMiaolingian of the North China Block (modified from Feng et al., 2002)
图 2 登封地区朱砂洞组综合柱状图及野外和显微镜照片
(a) gypsum karst breccia; (b) stromatolite and calcite dolomite; (c) laminated calcite dolomite, plane-polarized light (PPL) ; (d) flat-pebble conglomerate; (e) bimodal cross-bedding fine grained limestone; (f) fine grained limestone, PPL; (g) laminated calcite dolomite; (h) calcite dolomite, staining of alizarin red, PPL
Figure 2. Synthetical strata, field and microscopic photos of Zhushadong Formation in Dengfeng area
Fig.2
图 3 登封地区朱砂洞组Thalassinoides suevicus野外和显微镜照片
(a) large number of T.suevicus in the bedding plane, with gray filling, BI is 3; (b) large number of T.suevicus in the bedding plane, with earthy yellow filling, BI is 4; (c) T.suevicus in a vertical section, with dark gray filling, BI is 5; (d) T.suevicus in a vertical section, with earthy yellow filling, BI is 2; (e, f) limestone containing T.suevicus, PPL, the white arrows mark the T.suevicus
Figure 3. Field and microscopic photos of Thalassinoide suevicus from the Zhushadong Formation in the Dengfeng area
Fig.3
图 4 洛阳地区张夏组综合柱状图及野外和显微镜照片
(a) laminated limestone and mudstone with flat-pebble conglomerate; (b) stromatolite limestone; (c) bioturbated limestone with thrombolites; (d) thrombolites, PPL; (e) grainstone and bioturbated limestone; (f) bioturbated limestone, PPL; (g) thick-bedded oolitic limestone and bioturbated limestone; (h) oolitic limestone, PPL
Figure 4. Synthetical strata, field and microscopic photos of Zhangxia Formation in Luoyang area
Fig.4
图 5 洛阳地区张夏组Thalassinoides野外和显微镜照片
(a) T. horizontalis Type1 in the bedding plane indicated by the white dotted line, BI is 2; (b) T. horizontalis Type1 in three beds, indicated by the dotted line, different colors represent different beds of trace fossil, BI is 2; (c) T. horizontalis Type2 in the bedding plane, earthy yellow filling, BI is 3; (d) microscopic photo of T. horizontalis Type2, the filling material is coarse, part of it is iron-mineralized, and the surrounding rock is micrite calcite; (e) T.bacae in a vertical section, the vertical disturbance depth is large with localized branching, BI is 2; (f) T.bacae in a vertical section, BI is 3; (g) T.bacae in the bedding plane, BI is 3; (h) limestone containing T.bacae, the composition difference of burrow filling, lining, halo, and surrounding rock can be clearly seen, PPL
Figure 5. Field and microscopic photos of Thalassinoides from the Zhangxia Formation in the Luoyang area
Fig.5
图 6 鹤壁地区崮山组综合柱状图及野外和显微镜照片
(a) light gray limestone and grayish yellow mudstone interlayer; (b) conglomerate, micrite, and oolitic limestone; (c) tiny trace fossil Mycellia isp.; (d) trilobite debris on mudstone in the bedding plane; (e) dark grey thin-bedded micrite; (f) micrite and mudstone, PPL; (g, h) bioturbated limestone, grainstone, and microbialite
Figure 6. Synthetical strata, field and microscopic photos of the Gushan Formation in the Hebi area
Fig.6
图 7 鹤壁地区崮山组Thalassinoides bacae野外和显微镜照片
(a-c) T.bacae in the bedding plane; (d) T.bacae in a vertical section; (e, f) limestone containing T.bacae, the composition difference of burrow filling, lining, halo and surrounding rock can be clearly seen, PPL
Figure 7. Field and microscopic photos of Thalassinoidesbacae from the Gushan Formation in the Hebi area
Fig.7 -
[1] 冯增昭,陈继新,吴胜和. 1989. 华北地台早古生代岩相古地理[J]. 沉积学报,7(4):15-55. Feng Zengzhao, Chen Jixin, Wu Shenghe. 1989. Lithofacies paleogeography of Early Paleozoic of North China Platform[J]. Acta Sedimentologica Sinica, 7(4): 15-55. [2] 冯增昭,彭勇民,金振奎,等. 2002. 中国中寒武世岩相古地理[J]. 古地理学报,4(2):1-11. Feng Zengzhao, Peng Yongmin, Jin Zhenkui, et al. 2002. Lithofacies palaeogeography of the middle Cambrian in China[J]. Journal of Palaeogeography, 4(2): 1-11. [3] 冯增昭,彭勇民,金振奎,等. 2004. 中国寒武纪和奥陶纪岩相古地理[M]. 北京:石油工业出版社:1- 200. Feng Zengzhao, Peng Yongmin, Jin Zhenkui, et al. 2004.Lithofacies paleogeography of the Cambrian and Ordovician in China[M]. Beijing: Petroleum Industry Press: 1-200. [4] 李凯楠,王敏,齐永安,等.2019. 豫西寒武系苗岭统张夏组Thalassinoides bacae中莓状黄铁矿的成因及其生态学意义[J]. 古生物学报,58(4):445-455. Li Kainan, Wang Min, Qi Yong'an, et al. 2019. Framboidal pyrites in Thalassinoides bacae from the Zhangxia Formation (Cambrian Miaolingian series) in Henan pro-vince: Genesis and its ecological significance[J]. Acta Palaeontologica Sinica, 58(4): 445-455. [5] 罗茂,龚一鸣,张立军,等. 2021. 遗迹化石:探索生物与环境相互作用的重要信息载体[J]. 古生物学报,60(3):347-356. Luo Mao, Gong Yiming, Zhang Lijun, et al. 2021. Trace fossil: A significant agent for exploring organism-environment interactions[J]. Acta Palaeontologica Sinica, 60(3): 347-356. [6] 牟传龙,周恳恳,陈小炜,等. 2016. 中国岩相古地理图集(埃迪卡拉纪—志留纪)[M]. 北京:地质出版社:1- 154. Mou Chuanlong, Zhou Kenken, Chen Xiaowei, et al. 2016. Atlas of lithofacies and paleogeography of China (Ediacaran-Silurian)[M]. Beijing: Geological Publishing House: 1-154. [7] 裴放. 2000. 河南省华北型寒武纪生物地层单位划分与对比[J]. 河南地质,18(2):97-106. Pei Fang. 2000. Division and correlation of the North China type Cambrian biostratigraphic units of Henan province[J]. Henan Geology, 18(2): 97-106. [8] 裴放,张海清,阎国顺,等. 2008. 河南省地层古生物研究(第三分册)早古生代(华北型)[M]. 郑州:黄河水利出版社:1- 302. Pei Fang, Zhang Haiqing, Yan Guoshun, et al. 2008. The third Paleozoic study of stratigraphic paleontology in Henan province, Early Paleozoic (North China)[M]. Zhengzhou: Yellow River Water Conservancy Publishing House: 1-302. [9] 彭勇民,张荣强,陈霞,等. 2012. 四川盆地南部中下寒武统石膏岩的发现与油气勘探[J]. 成都理工大学学报(自然科学版),39(1):63-69. Peng Yongmin, Zhang Rongqiang, Chen Xia, et al. 2012. Dis-covery and significance of the middle-lower Cambrian gypsolith in the south of Sichuan Basin, China[J]. Journal of Chengdu University of Technology (Science & Technology Edition), 39(1): 63-69. [10] 齐永安,王敏,李妲,等. 2012. 寒武纪底质革命:从微生物席底到生物扰动混合底[J]. 河南理工大学学报(自然科学版),31(2):159-164. Qi Yong'an, Wang Min, Li Da, et al. 2012. Cambrian substrate revolution: From matgrounds to bioturbated mixgrounds[J]. Journal of Henan Polytechnic University (Natural Science), 31(2): 159-164. [11] 许晴旸,范若颖,龚一鸣. 2023. 海相遗迹化石对显生宙生物大辐射事件的响应[J]. 古地理学报,2023,25(2):431-450. Xu Qingyang, Fan Ruoying, Gong Yiming. 2023. Marine ichnofossils as a record of major biodiversification events in the Phanerozoic[J]. Journal of Palaeogeography, 25(2): 431-450. [12] 朱茂炎,杨爱华,袁金良,等. 2019. 中国寒武纪综合地层和时间框架[J]. 中国科学:地球科学,49(1):26-65. Zhu Maoyan, Yang Aihua, Yuan Jinliang, et al. 2019. Cambrian integrative stratigraphy and timescale of China[J]. Science China Earth Sciences, 49(1): 26-65. [13] Bromley R G, Ekdale A A. 1984. Trace fossil preservation in flint in the European chalk[J]. Journal of Paleontology, 58(2): 298-311. [14] Buatois L A, Mángano M G. 2004. Animal-substrate interactions in freshwater environments: Applications of ichnology in facies and sequence stratigraphic analysis of fluvio-lacustrine successions[J]. Geological Society, London, Special Publications, 228(1): 311-333. [15] Buatois L A, Mángano M G. 2011. Ichnology: Organism-substrate interactions in space and time[M]. Cambridge: Cambridge University Press: 1-358. [16] Carmona N B, Buatois L A, Mángano M G. 2004. The trace fossil record of burrowing decapod crustaceans: Evaluating evolutionary radiations and behavioural convergence[J]. Fossils and Strata, 51: 141-153. [17] Droser M L, Bottjer D J. 1986. A semiquantitative field classification of ichnofabric[J]. Journal of Sedimentary Research, 56(4): 558-559. [18] Dworschak P C, Rodrigues S D A. 1997. A modern analogue for the trace fossil Gyrolithes: Burrows of the thalassinidean shrimp Axianassa australis [J]. Lethaia, 30(1): 41-52. [19] Ekdale A A, Bromley R G. 2003. Paleoethologic interpretation of complex Thalassinoides in shallow-marine limestones, Lower Ordovician, southern Sweden[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 192(1/2/3/4): 221-227. [20] Ekdale A A, Lamond R E. 2003. Behavioral cladistics of trace fossils: Evolution of derived trace-making skills[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 192(1/2/3/4): 335-343. [21] El-Sabbagh A, El-Hedeny M, Al Farraj S. 2017. Thalassinoides in the Middle Miocene succession at Siwa Oasis, northwestern Egypt[J]. Proceedings of the Geologists' Association, 128(2): 222-233. [22] Erwin D H. 2008. Macroevolution of ecosystem engineering, niche construction and diversity[J]. Trends in Ecology & Evolution, 23(6): 304-310. [23] Feng X Q, Chen Z Q, Benton M J, et al. 2019. A diverse trackway-dominated marine ichnoassemblage from the Lower Triassic in the northern Paleotethys: Ichnology and implications for biotic reco-very[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 519: 124-140. [24] Griffis R B, Suchanek T H.1991. A model of burrow architecture and trophic modes in thalassinidean shrimp (Decapoda: Thalassinidea)[J]. Marine Ecology Progress Series, 79: 171-183. [25] Jin J S, Harper D A T, Cocks L R M, et al. 2013. Precisely locating the Ordovician equator in Laurentia[J]. Geology, 41(2): 107-110. [26] Jin J S, Harper D A T, Rasmussen J A, et al. 2012. Late Ordovician massive-bedded Thalassinoides ichnofacies along the palaeoequator of Laurentia[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 367/368: 73-88. [27] Joseph J K, Patel S J, Bhatt N Y. 2012. Trace fossil assemblages in mixed siliciclastic-carbonate sediments of the Kaladongar Formation (Middle Jurassic), Patcham Island, Kachchh, western India[J]. Journal of the Geological Society of India, 80(2): 189-214. [28] Knaust D. 2021. Balanoglossites-burrowed firmgrounds:The most common ichnofabric on earth?[J]. Earth-Science Reviews, 220: 103747. [29] Knaust D. 2024. The trace fossil Thalassinoides paradoxicus Kennedy, 1967 revisited from its type locality (Albian-Cenomanian chalk, SE England)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 634: 111913. [30] Knaust D, Bromley R G. 2012. Trace fossils as indicators of sedimentary environments[M]. Amsterdam: Elsevier Science: 1-924. [31] Leonowicz P. 2015. Ichnofabrics of shallow-marine mudstone, the result of changing environmental conditions: An example from the Middle Jurassic ore-bearing clay from southern Poland[J]. Facies, 61(3): 11. [32] Liu B C, Qi Y A, Wang M, et al. 2024. Thalassinoides ichnofabrics from the Zhangxia Formation (Cambiran Miaolingian series) in Henan province: Bioturbators as Ecosystem engineers[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 640: 112103. [33] Marusin V V, Kochnev B B, Izokh O P. 2022. Thalassinoides and Olenichnus in the Terreneuvian carbonates of the Igarka uplift, NW Siberian Platform[J]. Lethaia, 55(2): 1-17. [34] McIlroy D, Logan G A. 1999. The impact of bioturbation on infaunal ecology and evolution during the Proterozoic-Cambrian transition[J]. Palaios, 14(1): 58-72. [35] Rodríguez-Tovar F J, Miguez-Salas O, Duarte L V. 2017. Toarcian Oceanic Anoxic Event induced unusual behaviour and palaeobiological changes in Thalassinoides tracemakers[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 485: 46-56. [36] Scholle P A, Ulmer-Scholle D S. 2003. A color guide to the petrography carbonate rocks: Grains, textures, porosity, diagenesis[M]. Tulsa: American Association of Petroleum Geologists: 1-459. [37] Seilacher A. 2007. Trace fossil analysis[M]. Berlin: Springer: 1-226. [38] Sharafi M, Ashuri M, Mahboubi A, et al. 2012. Stratigraphic application of Thalassinoides ichnofabric in delineating sequence stratigraphic surfaces (Mid-Cretaceous), Kopet-Dagh Basin, northeastern Iran[J]. Palaeoworld, 21(3/4): 202-216. [39] Sheehan P M, Schiefelbein D R J. 1984. The trace fossil Thalassinoides from the Upper Ordovician of the eastern Great Basin: Deep burrowing in the Early Paleozoic[J]. Journal of Paleontology, 58(2): 440-447. [40] Solan M, Kennedy R. 2002. Observation and quantification of in situ animal-sediment relations using time-lapse sediment profile imagery (t-SPI)[J]. Marine Ecology Progress Series, 228: 179-191. [41] Taylor A, Goldring R, Gowland S. 2003. Analysis and application of ichnofabrics[J]. Earth-Science Reviews, 60(3/4): 227-259. [42] Torsvik T H, Cocks L R M. 2019. The integration of palaeomagnetism, the geological record and mantle tomography in the location of ancient continents[J]. Geological Magazine, 156(2): 242-260. [43] Uchman A, Wetzel A. 2012. Deep-sea fans[J]. Developments in Sedimentology, 64: 643-671. [44] Wang M, Li K N, Yang W T, et al. 2019. The trace fossil Thalassinoides bacae in the Cambrian Zhangxia Formation (Miaolingian series) of North China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 534: 109333. [45] Wetzel A, Rodríguez-Tovar F J. 2021. Preface: Ichnology, trace fossils and depositional environments[J]. Palaeogeography, Palaeo-climatology, Palaeoecology, 565: 110220. [46] Zhang X L, Wang Y, Wang Y X. 2024. Widespread Thalassinoides facies from the Upper Silurian of South China: A case study from the northwestern Hunan province, Middle to Upper Yangtze Block[J]. Palaeoworld, 33(6): 1545-1563. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 365
- HTML全文浏览量: 58
- PDF下载量: 169
- 被引次数: 0

