

 下载:
下载:







-
现代沉积作用是物理、化学以及生物三者相互作用的产物(胡斌等,2012;王媛媛等,2019)。新遗迹学研究通过观察现代生物的造迹活动了解其活动规律、遗迹特点和分布特征等(胡斌等,2012;王媛媛等,2019,2024;Nascimento et al.,2021;王翠等,2023),为潜穴和它们的生产者以及它们的生活环境条件之间的联系提供了证据(Wang et al.,2019b)。通过对现代生物遗迹的详细研究,可以更好地了解现代和古代沉积中潜穴形态与生物行为之间的关系(Counts and Hasiotis,2009),对古环境、古生态、古土壤的解释有一定的启发性(Brake et al.,2002;Hasiotis,2004;Hembree and Hasiotis,2006;Kraus and Hasiotis,2006;Counts and Hasiotis,2009;Abdel‑Fattah,2019;Toniolo et al.,2021)。
潮坪作为海陆过渡的重要地带,其复杂多变的环境孕育了丰富多样的生物遗迹。之前对潮坪环境的研究大多局限于物理沉积特征,如粒度分析(刘仲衡等,1985)、潮汐动力(范代读和李从先,2000)、沉积韵律(李铁松和李从先,1995)、层理特征(Miller and Eriksson,1997)等。随着新遗迹学迅速发展,国内外学者对潮坪环境中的现代生物遗迹也进行了较为深入的研究。王媛媛团队使用高精度的三维重构技术和先进的分析手段讨论了黄河三角洲潮坪环境中生物遗迹与物理、化学条件之间的响应关系(王媛媛等,2019;Wang et al.,2019a,2019b;王翠等,2023)。Swinbanks等学者研究了加拿大弗雷泽河三角洲边界湾潮坪生物遗迹的分布以及不同亚环境中独特的生物沉积结构(Swinbanks and Murray,1981;Swinbanks and Luternauer,1987)。Lee and Koh(1994)利用树脂浇注法等,研究了韩国西海岸泥滩主要底栖动物遗迹的形态和沉积构造。然而,潮坪中潮汐沉积物特有的生物扰动遗迹学模式仍未被系统提出。只有少数学者尝试建立潮汐环境的遗迹学模式(Mángano and Buatois,2004;McIlroy,2004),认为潮汐遗迹模式类似于半咸水环境的遗迹模式即Cruziana遗迹相模式和Skolithos遗迹相模式或者是Cruziana-Skolithos混合相模式,并在中生代和新生代的现代和古代环境中已经得到了很好的验证(Pemberton et al.,1982;MacEachern et al.,1999;Gingras et al.,2007)。但是专门针对潮汐沉积过程中造迹生物的造迹行为方面的研究较少,如潮汐能量变换期间,造迹生物对潜穴的重建,或者在变化的潮汐底质中的机会主义殖居行为等(Taylor et al.,2003;Bann et al.,2004;Dashtgard et al.,2008)。因此,研究潮坪环境中现代生物遗迹对潮汐—遗迹学模型的建立和沉积环境解释有着重要意义(Taylor et al.,2003;Bann et al.,2004;Kumar,2017;Lan et al.,2021)。
本次研究对珠江三角洲潮坪环境中现代生物的种类及其行为习性和生物营造的遗迹进行了详细的描述,介绍了不同微环境中现代生物遗迹的组成和分布并补充了现代潮坪沉积环境中的生物学信息,以期对古潮坪沉积环境及潮坪遗迹相进行类比提供现代沉积资料。
-
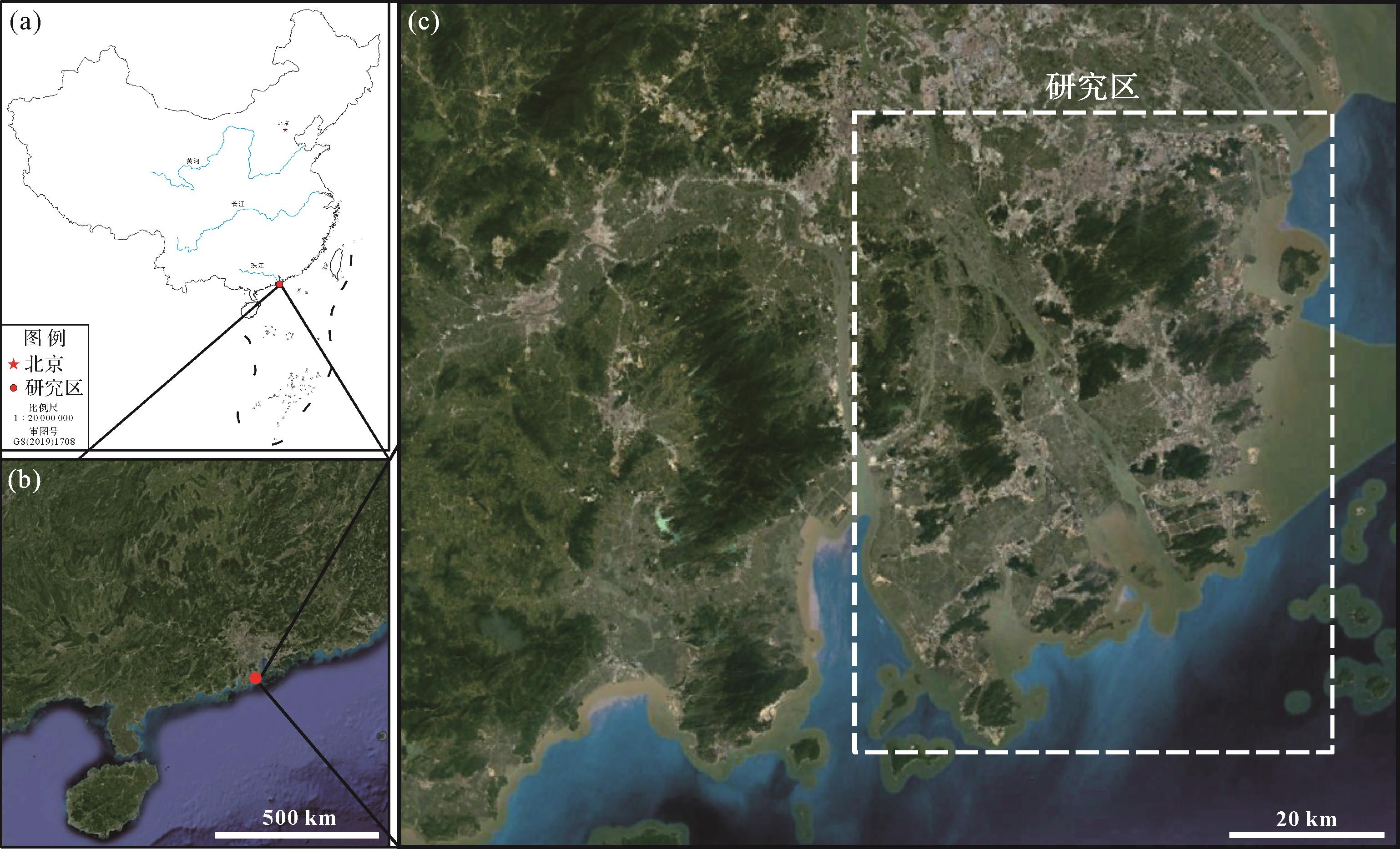
研究区属于珠江三角洲的海岸潮坪环境(图1)。珠江三角洲是中国第二大河流三角洲(Chan et al.,2021),由东江、西江和北江等多条河流在珠江河口湾内堆积而成(韦惺和吴超羽,2018;张绍轩等,2020;时硕等,2022),区域内水文受到珠江径流和南海海潮相互作用的影响(Chan et al.,2021)。

图 1 研究区位置图
Figure 1. Location map of the study area
东江、西江和北江是珠江三角洲的三条主要支流(Zhang et al.,2008;Li et al.,2016),三条河流占珠江总流量的89.6%,西江年径流量占珠江总流量的70.8%(Geng et al.,2015)。珠江年径流量约为3 360×108 m3,高流量期(4—9月)径流量通常占年径流量的80%(Geng et al.,2015)。三角洲属亚热带季风气候,年平均气温21 ℃~23 ℃,年平均降水量1 600~2 600 mm(Zhang et al.,2008;Li et al.,2016;Zhang et al.,2019),4月至9月的雨季约占全年降雨量的80%(Chan et al.,2021)。南海北部陆架以不规则的半日潮为主,潮差为2~4 m(Li et al.,2016)。在总泥沙排放中,20%沉积在三角洲,80%进入海洋,从而导致珠江口地区以每年40 m的速度向海延伸(Weng,2007)。
本次研究选取珠江三角洲沿岸地区的潮坪环境,潮上带和潮间带生物扰动明显,造迹生物的种类及其遗迹丰富,为主要研究区。
-
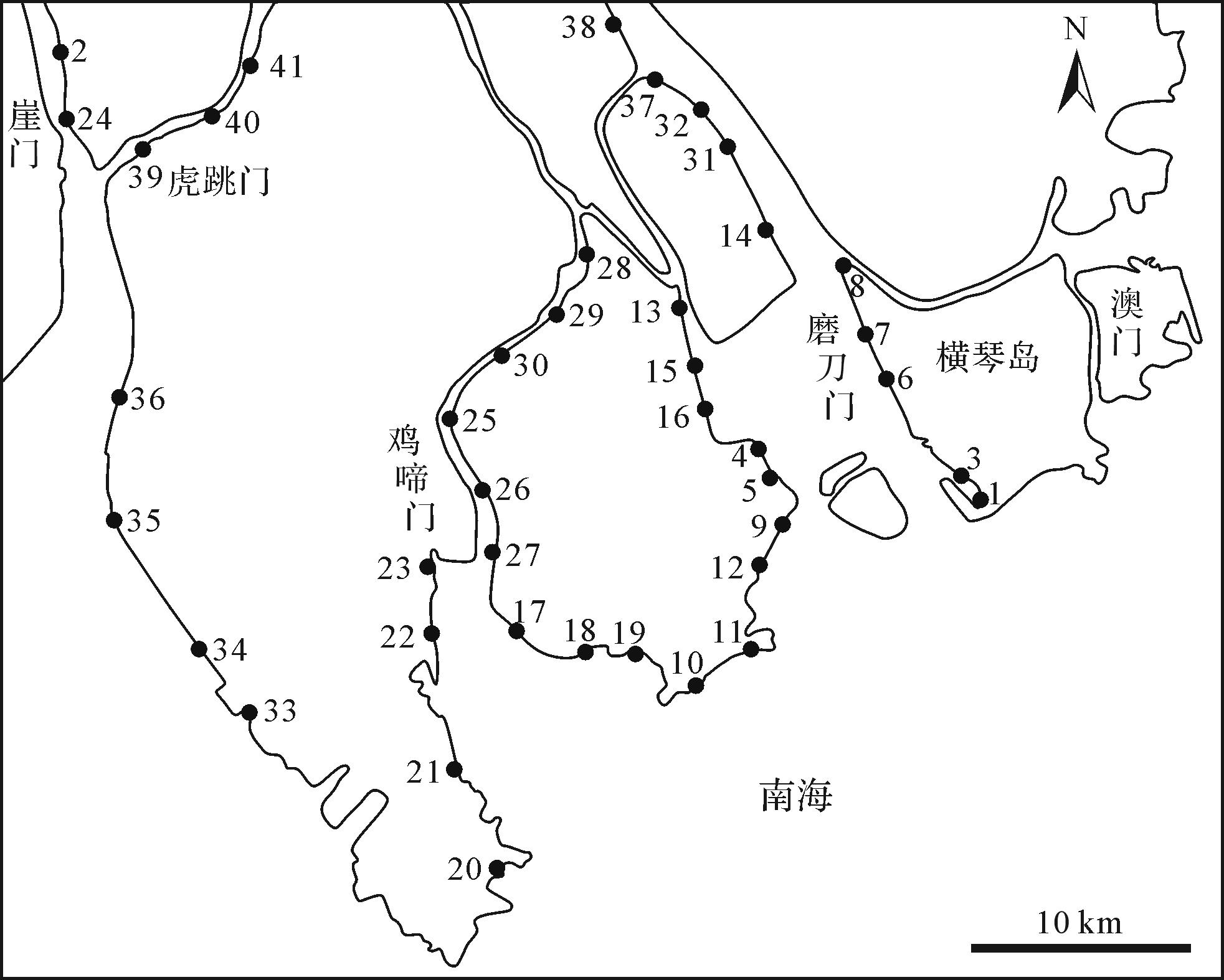
野外踏勘时共在崖门、虎跳门、鸡啼门和磨刀门四大口门水道及其周围海岸的潮坪选取采样点41个。使用GPS确定点位的经纬度坐标,并在Google Earth Pro进行标记后绘制(图2)。对各个点位的造迹生物及其遗迹进行了详细的观察和描述,同时使用以聚氯乙烯树脂为主要原料的PVC管(直径为7.5 cm,高度为15 cm)在部分点位对形态保存完好且有代表性的生物潜穴进行取心。对取心后的PVC 管,用防水胶带密封其上下口,以防止泥土和水体渗漏。在人为运输至进行扫描的过程中,要把PVC管固定好,始终确保 PVC 管处于潜穴口朝上的直立稳定状态,尽可能避免其内部遗迹原有形态发生变形。

图 2 四大口门潮坪采样点分布图
Figure 2. Distribution map of sampling points in tidal flats of the four major estuaries
-
使用纳米X射线数字岩心分析系统对采取的岩心样品进行扫描,通过处理扫描取得的切片数据还原生物营造的层内潜穴形态。扫描仪器参数如下:型号是Nanotom S,电压是180 kV,功率是15 W,细节检测能力高达200 nm。CT扫描实验在中国科学院南京土壤研究所完成。
-
三维重构即对CT扫描获取的CT图像数据(体数据)进行三维可视化处理,包括体数据三维渲染、灰度值分割、定向数字虚拟切面、动画制作和图片的保存等。处理过程中使用到ImageJ和VG Studio Max 3.0两款软件。三维重构过程在中国科学院南京古生物地质研究所X射线断层扫描实验室完成。
具体处理方式如下:(1)将扫描获取的切片图导入至ImageJ,观察生物潜穴是否清晰可见,将效果好的孔隙通过ImageJ进行提取,效果不好的进行降位(图片的位深度由16 bit降至8 bit)。(2)将在ImageJ中处理过的图片导入VG Studio 3.0再次处理,最后对制作完成的生物潜穴进行渲染和动画的制作,将图片和视频保存。
-
珠江三角洲潮坪环境现代生物遗迹主要集中在潮上带和潮间带,潮间带生物遗迹分异度和丰度最高。潮上带采样点较少,只有采样点1和采样点14-15。潮间带根据其不同的沉积底质划分为泥坪(采样点1-2,6,8,13,21-22,27,29-33,36-38,40-41)、混合坪(3-5,7,10-11,17,23-26,28,34,39)和沙坪(9,12,14-16,18-20,35)。
-
潮上带处于平均高潮线以上,只有在台风和风暴等恶劣天气影响下的特大高潮才会被淹没。该区域的沉积底质较潮间带和潮下带偏硬,主要造迹生物包括海蟑螂和弧边招潮蟹等。弧边招潮蟹在潮上带营造的潜穴形态单一,有点坑状的足辙迹等,与潮间带的泥坪和混合坪中的部分弧边招潮蟹的遗迹类似,和潮间带生物遗迹统一描述。
-
潮间带处于平均高潮线和平均低潮线之间,涨潮时被海水覆盖,退潮时裸露。该区域的沉积底质一般较为松软,可分为泥坪、混合坪和沙坪。潮间带生物种类丰富,弧边招潮蟹、纠结招潮蟹和清白招潮蟹等招潮蟹属的蟹类居多,招潮蟹属的螃蟹在研究区内营造的遗迹类似,此处仅以弧边招潮蟹为例进行描述。此外,还有一些其他种类的螃蟹和双齿围沙蚕、弹涂鱼、螺类以及鸟类等。
-
1) 弧边招潮蟹Uca arcuata营造的遗迹
弧边招潮蟹Uca arcuata为沙蟹科招潮蟹属,主要分布在潮上带以及潮间带的泥坪和混合坪。泥坪和混合坪环境中数量最多,生物遗迹也更丰富。弧边招潮蟹营造的遗迹主要为层面的进食迹、排泄迹、足辙迹和层内的居住潜穴。弧边招潮蟹的活动跟随潮水涨落的规律,涨潮时在潜穴内休息,退潮时在潮坪上活动和取食。弧边招潮蟹以沉积物中的有机物为食,并将不能利用的残渣置于地面形成排泄迹(图3a),其在潮坪上活动时常留下点坑状的足辙迹(图3b,c)。弧边招潮蟹的潜穴形态复杂多样,层内潜穴与层面垂直或斜交,层面的潜穴口呈近圆形(图3c),还发现土丘状(图3d)和烟囱状(图3e)的潜穴口。土丘状和烟囱状的潜穴口可能是雌蟹用来保护自己或雄蟹用于求偶。通过观察CT扫描和三维重构,恢复了弧边招潮蟹的层内3D潜穴形态,其形态为简单的I形和弧形(图3f)。

图 3 弧边招潮蟹营造的遗迹
Figure 3. Trace produced by Uca arcuata
2) 弹涂鱼Periophthalmus cantonensis营造的遗迹
弹涂鱼Periophthalmus cantonensis为虾虎鱼科弹涂鱼属,主要分布在潮间带的泥坪环境,在混合坪和沙坪中也有发现,未在沙坪中营造潜穴。弹涂鱼营造的遗迹主要是爬行迹和居住迹。弹涂鱼营底栖生活,潜穴常建在底质为淤泥或泥沙的滩涂上,退潮时会外出运动觅食,其中有其营造的丰富遗迹。弹涂鱼能在滩涂中运动归功于其发达的胸鳍,胸鳍前后自如运动,拖动身体向前。运动过程中,弹涂鱼胸鳍的压痕留下明显的纹理(图4a),身体拖动时会留下犁沟状的拖痕,犁沟两侧是对称的胸鳍压痕(图4b)。不同沉积底质中爬行迹保存也有区别,爬行迹在泥质沉积物表面比在砂质沉积物表面更清晰,也更容易保存(图4b,c)。弹涂鱼潜穴是其觅食、居住和产卵的场所,受到惊吓时会迅速逃回潜穴(曾洪根,2019)。潜穴一般有一至两个开口,双开口能起到更好的保护作用。潜穴口通常有爬行迹(图4c),部分潜穴口会有隆起(图4d)。弹涂鱼双开口的层内潜穴一般为Y形(图4e)或U形,单开口的为I形,穴口与层面倾斜。

图 4 弹涂鱼营造的遗迹
Figure 4. Trace produced by Periophthalmus cantonensis
3) 无齿东方相手蟹Orisarma dehaani营造的遗迹
无齿东方相手蟹Orisarma dehaani为方蟹总科相手蟹属,主要分布在口门附近潮间带的泥坪环境,盐度较低,靠海沿岸地区的潮坪中没有发现其生活的痕迹。无齿东方相手蟹营造的遗迹主要为足辙迹和居住迹。无齿东方相手蟹幼蟹常在较为松软的泥质中掘穴和觅食(图5a),成蟹的潜穴筑在较为坚硬的粉砂质底质中(图5b)或隐藏于石块间(图4c)。无齿东方相手蟹的潜穴呈近圆形,部分潜穴有围墙状的轻微隆起(图5d)或泥浆散落在穴口附近。穴口倾斜于层面,附近有凌乱的足辙迹(图5e)。无齿东方相手蟹营造的层内潜穴多样,常见的有垂直或稍倾斜于层面的I形以及有两个穴口的Y形潜穴(图5f)。

图 5 无齿东方相手蟹营造的遗迹
Figure 5. Trace produced by Chiromantes dehaani
-
1) 双齿围沙蚕Perinereis aibuhitensis营造的遗迹
双齿围沙蚕Perinereis aibuhitensis为沙蚕科围沙蚕属,主要分布在潮间带的混合坪环境,在泥坪环境中也有发现。双齿围沙蚕幼虫以浮游生物为食,成虫以腐殖质为食。双齿围沙蚕营造的遗迹主要为居住迹和排泄迹,未发现其在层面营造的痕迹。沙蚕潜穴口一般为近圆形,直径2 mm左右。沙蚕会摄取层面上的藻类和有机质,在层面游走时会排出细条状的排泄物(图6a),分布于潜穴周围。通过观察岩心剖面(图6b,c)以及CT扫描和三维重构获得的沙蚕层内潜穴三维形态(图6d),可以看出沙蚕层内潜穴十分复杂,形态主要为I形、U形和Y形。有时可以观察到沙蚕和螃蟹共生现象,这是因为沙蚕在层内觅食沉积物中的有机质时可能会误入螃蟹潜穴,沙蚕也是螃蟹等甲壳类动物的重要食物。

图 6 双齿围沙蚕营造的遗迹
Figure 6. Trace produced by Perinereis aibuhitensis
2) 海蟑螂Ligia oceanica营造的遗迹
海蟑螂Ligia oceanica为海蟑螂科海蟑螂属,主要分布在潮间带的混合坪和潮上带环境。海蟑螂一般成群活动,反应灵敏,爬行迅速,为杂食性,多以藻类为食。海蟑螂幼虫常在潮间带的混合坪中营造潜穴生活,成虫一般生活在高潮间带或潮上带海浪浸湿不到的岩石缝中。幼虫营造的居住潜穴多呈近圆形或椭圆形,潜穴十分密集(图7a)。层内潜穴形态十分多样,其潜穴剖面上有Y形、U形和垂直于层面的I形潜穴(图7b)。

图 7 海蟑螂、幡条蜑螺、黄口蜑螺营造的遗迹
Figure 7. Traces produced by Ligia oceanica, Vittina coromandeliana, Neritina pulligera
3) 幡条蜑螺Vittina coromandeliana营造的遗迹
幡条蜑螺Vittina coromandeliana为蜑螺科饰纹蜑螺属,在潮间带泥坪和混合坪环境中均有出现。幡条蜑螺喜食藻类,壳体呈水滴形,表面光滑。幡条蜑螺营造的遗迹主要为爬行迹,其通常在沉积物表面和附近的礁石上缓慢爬行,形成不规则的爬行迹(图7c)。
4) 黄口蜑螺Neritina pulligera营造的遗迹
黄口蜑螺Neritina pulligera为蜑螺科游螺属,分布在潮间带混合坪环境。黄口蜑螺在退潮后的层面上爬行,数量较少,营造的主要是爬行迹。混合坪中发现的黄口蜑螺壳体大小在1.5 cm左右,移动时在层面形成连续的爬行迹。爬行迹宽约1 cm,长度不定,爬行迹有断断续续的弧状纹理(图7d)。
-
中华沙蟹Ocypode cordimand为沙蟹科沙蟹属,分布在潮间带沙坪环境。中华沙蟹营造的遗迹主要为进食迹、排泄迹和居住迹。中华沙蟹主要以腐殖质和沉积物中的有机质为食,其通常在洞穴周围觅食,将沉积物中无用的部分过滤后放置在层面上(图8a)。沙蟹堆积的拟粪球粒无规则地分布在其洞穴周围,呈星射状(图8a)、鸟尾状(图8b)和扇形(图8c)等,但通常会留一条逃跑路线。沙蟹常成群出现对大片沙滩进行改造(图8d),场面极其壮观。沙蟹的潜穴多筑在平坦的沙滩上,呈近圆形,直径较小,介于5~15 mm,其层内潜穴多为垂直于平面的I形(图8e)。

图 8 中华沙蟹营造的遗迹
Figure 8. Trace produced by Ocypode cordimand
-
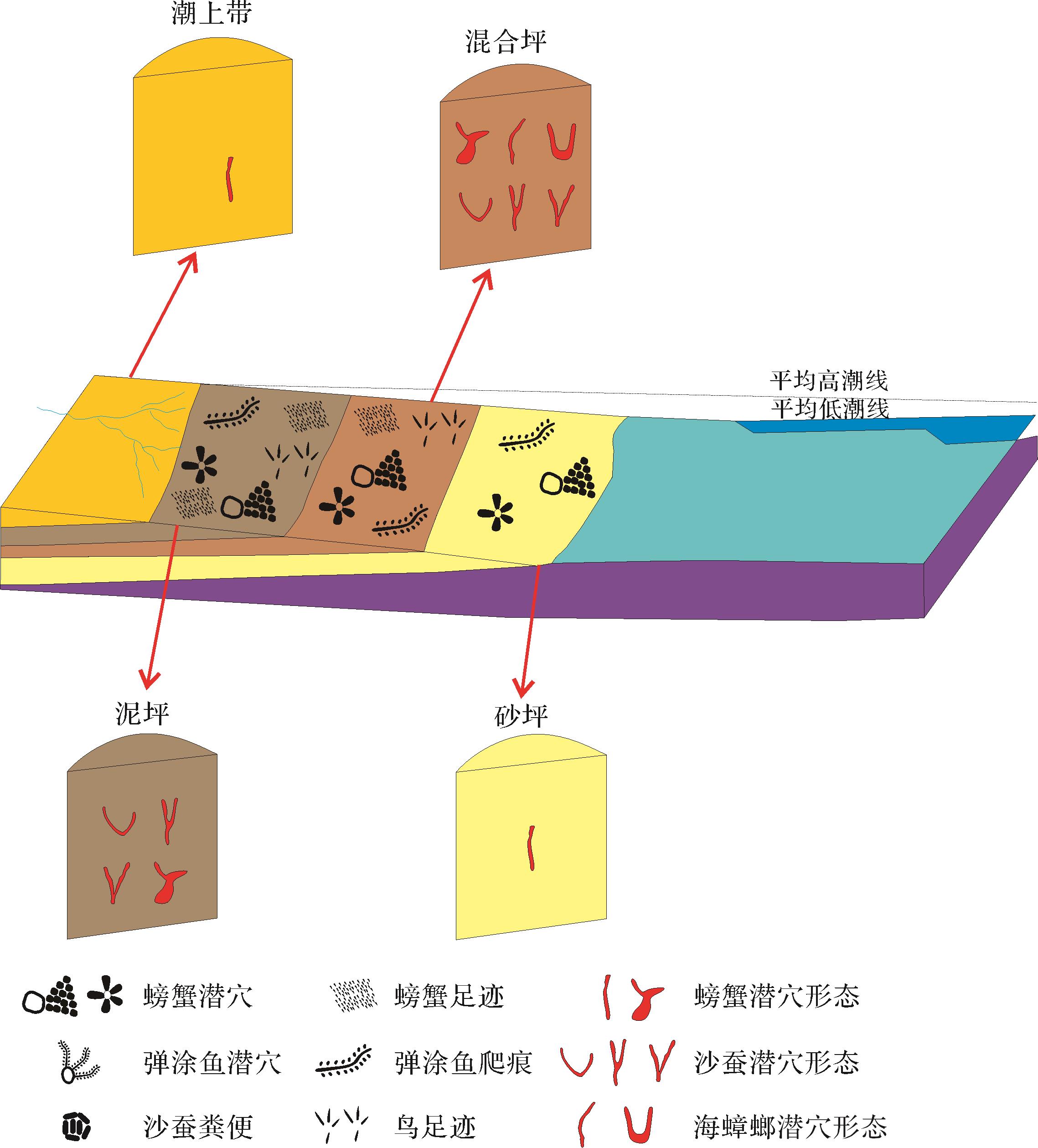
潮下带位于平均低潮线以下,受潮汐和波浪的共同作用,水动力较强。该环境生物遗迹较少且不便于观察,此处不作描述(图9)。

图 9 珠江三角洲潮坪环境现代生物遗迹组成及分布模式图
Figure 9. Composition and distribution mode of modern biological traces in tidal flat environment of the Pearl River Delta
-
在珠江三角洲潮坪环境中发现了多种造迹生物和不同的生物遗迹组合,这是因为造迹生物及其遗迹的分布与潮坪环境中的微环境密切相关。潮坪沉积物中丰富的资源可能会使某些特征遗迹属占据优势地位。那些能够快速占据沉积物底层环境(例如Planolites和Thalassinoides),或者从沉积物—水界面上获取资源的生物行为(例如Skolithos、Arenicolites和双壳类产生的Cylindrichnus),使得这些遗迹最终得以保存于潮坪沉积物之中,成为潮坪沉积的遗迹群落(Gingras et al.,1999)。此外,在不同的沉积环境中,沉积物特征、营养物质来源和水动力条件等存在差异,也会导致造迹生物及其遗迹在丰度和分异度上有所不同,可分为以下两种遗迹组合模式。
潮上带为低能高潮区,正常潮汐作用下不能到达,故该区域常露出水面,沉积物粒度较粗,底质含水量较低,食物稀缺,生物遗迹种类和数量较少。潮上带的生物遗迹组合主要为海蟑螂Ligia oceanica的Y形、U形和垂直于层面的I形居住潜穴以及少量的弧边招潮蟹Uca arcuata的居住潜穴。前者的潜穴丰度较高,并且常出现在岩石缝中,这一区域的环境较为干燥,且受到潮汐作用较小,能够为海蟑螂提供较为稳定的栖息地。弧边招潮蟹一般属于潮间带生物,在潮上带发现的原因可能是潮间带在潮汐的作用下,能够提供一定的水流速度和方向(Drolet et al.,2012),影响了弧边招潮蟹的迁移模式,增强了它们的扩散能力(Talke and Stacey,2003),使其在潮上带也有出现。
潮间带处于海洋与陆地之间的过渡地带,沉积特征多样,沉积物含有泥质、砂质以及泥砂混合质等多种类型,这使得潮间带能够为不同的生物提供多样化的栖息和觅食空间。水动力相比潮上带较强,潮汐和波浪的动力又为造迹生物带来了充足的营养物质,有机物含量相对较高(Carter and Heath,1975),造迹生物及其遗迹的丰度和分异度较高。在潮间带中的生物遗迹主要有弧边招潮蟹Uca arcuata的层面进食迹、排泄迹、足辙迹和层内I形、弧形居住潜穴;弹涂鱼Periophthalmus cantonensis层面爬行迹和层内Y形、U形、I形居住潜穴;无齿东方相手蟹Orisarma dehaani层面足辙迹和层内I形、Y形居住潜穴;双齿围沙蚕Perinereis aibuhitensis层面排泄迹和层内I形、U形、Y形居住潜穴;海蟑螂Ligia oceanica层内Y形、U形、I形居住潜穴;幡条蜑螺Vittina coromandeliana和黄口蜑螺Neritina pulligera的爬行迹;中华沙蟹Ocypode cordimand层面进食迹、排泄迹和层内I形居住迹。其中弹涂鱼和双齿围沙蚕在潮间带的泥坪和混合坪均有发现,这是因为潮间带中的泥坪和混合坪均存在较为松软的泥质或砂质沉积底质,能够让弹涂鱼更好地建造潜穴,并且潮间带的泥质和砂质底质中含有较为丰富的有机物碎屑和微生物,这些均是弹涂鱼所需要的营养来源(张其永等,2006)。双齿围沙蚕偏好于在低潮区、中潮区和高潮区的环境中生存,有较强的适应能力(孟珊等,2020)。
-
珠江三角洲潮坪环境中现代生物遗迹在潮上带和潮间带分布居多,潮间带是主要的分布环境。生物遗迹主要包括层面遗迹和层内遗迹,造迹生物有螃蟹、弹涂鱼、海蟑螂、沙蚕和螺类,其中以螃蟹属为造迹生物的遗迹广为分布。层面遗迹主要包括进食迹、排泄迹、足辙迹等;层内遗迹以居住潜穴为主,形态多样,包括I形、U形和Y形。
此外,研究区内的现代生物遗迹丰度和分异度受水动力特征、粒度特征、食物来源等多种因素的影响。潮间带食物来源最为丰富,且水动力条件适中,再加上波浪和潮汐共同作用,为生物提供了多样的生存空间,因此该环境中生物遗迹丰度和分异度高于潮上带和潮下带。由此可反映出生物对潮坪环境的适应性。
Composition and Distribution of Modern Biological Traces in the Tidal Flat Environments of the Pearl River Delta
-
摘要: 目的 生物对其生活环境的反应极其灵敏,其营造的遗迹是生物与环境相互作用的结果,在一定程度上反映了沉积环境特征。生物沉积结构是了解现代海岸沉积环境中的生态学及沉积过程的有力工具。 方法 通过野外观察取样和三维重构等方法对珠江三角洲潮坪环境中现代生物遗迹的组成和分布特征进行了研究。 结果 潮上带、潮间带和潮下带生物分布差异较大,生物遗迹主要分布在潮间带。潮上带主要有海蟑螂和弧边招潮蟹,营造的遗迹主要有弧边招潮蟹的层面进食迹、排泄迹、足辙迹和层内I形居住潜穴。潮间带泥坪环境中的生物遗迹主要有弧边招潮蟹的层面进食迹、排泄迹、足辙迹和层内I形居住潜穴;弹涂鱼层面爬行迹和层内Y形、U形、I形居住潜穴;无齿东方相手蟹层面足辙迹和层内I形、Y形居住潜穴。潮间带混合坪环境中的生物遗迹主要有双齿围沙蚕层面排泄迹和层内I形、U形、Y形居住潜穴;海蟑螂层内Y形、U形、I形居住潜穴;幡条蜑螺和黄口蜑螺的爬行迹。潮间带沙坪环境中的生物遗迹主要有中华沙蟹层面进食迹、排泄迹和层内I形居住迹。潮下带环境不便观察。 结论 现代珠江三角洲潮间带生物遗迹的研究工作,不仅对珠江三角洲潮坪环境的现代沉积学资料进行了补充,也对古遗迹学和古环境重构提供了现代实证。Abstract: Objective Organisms are extremely responsive to the environments in which they live, and the traces they create are the result of interactions between organisms and their environment. To a certain extent, the traces reflect the properties of the depositional environment. Biological sedimentary structures are a powerful tool in understanding the ecology and depositional processes of modern coastal sedimentary environments. Methods The composition and distribution of modern biological traces in the tidal flat environments of the Pearl River Delta were studied by field observations and 3D reconstruction. Results The key understanding obtained by the study was that the distribution of organisms in the supratidal, intertidal and subtidal varies greatly, and the biological traces are mainly distributed in the intertidal zone. In the supratidal zone, mainly Ligia oceanica and Uca arcuata are present. The main traces are Uca arcuata level feeding traces, excretion traces, trackways, and intra-layer I-shaped dwelling burrows. Biological traces in the intertidal mud flat environment mainly include feeding, excretion, trackways and I-shaped dwelling burrows of Uca arcuata, crawling traces and Y-, U- and I-shaped dwelling burrows of Periophthalmus cantonensis, and trackways and I- and Y-shaped dwelling burrows of Orisarma dehaani. Biological traces in the mixed intertidal flat environment mainly include excretion traces and intra-layer I-, U-, and Y-shaped dwelling burrows of Perinereis aibuhitensis, Y-, U- and I-shaped dwelling burrows of Ligia oceanica, and crawling traces of Vittina coromandeliana and Neritina pulligera. Biological traces in the sandy intertidal environment include feeding and excretion traces of Ocypode cordimand and intra-layer I-shaped dwelling traces. The subtidal environment was not accessible for observation and no biological traces were found. Conclusions The study of modern intertidal biological traces in the Pearl River Delta supplements modern sedimentological data on tidal flat environments in the Pearl River Delta, and provides modern empirical evidence for palaeoichnology and palaeoenvironmental reconstruction.
-
Key words:
- biological traces /
- ichnology /
- tidal flat /
- Pearl River Delta
-
图 1 研究区位置图
(a) location of the study area, http://bzdt.ch.mnr.gov.cn/index.html from standard map service system of China; (b) study area bitmap (Google Earth Pro); (c) Pearl River Delta (Google Earth Pro)
Figure 1. Location map of the study area
Fig.1
图 2 四大口门潮坪采样点分布图
Figure 2. Distribution map of sampling points in tidal flats of the four major estuaries
图 7 海蟑螂、幡条蜑螺、黄口蜑螺营造的遗迹
Figure 7. Traces produced by Ligia oceanica, Vittina coromandeliana, Neritina pulligera
-
[1] 范代读,李从先. 2000. 长江三角洲泥质潮坪沉积的韵律性及保存率[J]. 海洋通报,19(6):34-41. Fan Daidu, Li Congxian. 2000. Lamination and preservation rate of mudflat deposition on the Changjiang Delta[J]. Marine Science Bulletin, 19(6): 34-41. [2] 胡斌,王媛媛,张璐,等. 2012. 黄河中下游焦作区段现代边滩沉积中的生物遗迹[J]. 古地理学报,14(5):628-638. Hu Bin, Wang Yuanyuan, Zhang Lu, et al. 2012. Biogenic traces in modern point bar deposits of the middle-lower reaches of Yellow River in Jiaozuo area, Henan province[J]. Journal of Palaeogeography, 2012, 14(5): 628-638. [3] 李铁松,李从先. 1995. 潮坪沉积韵律与沉积间断[J]. 同济大学学报,23(1):53-58. Li Tiesong, Li Congxian. 1995. Tidal flat depositional rhythm and diastem[J]. Journal of Tongji University, 23(1): 53-58. [4] 刘仲衡,吴锦秀,于永杰,等. 1985. 利用粒度资料浅析黄河三角洲潮坪沉积环境[J]. 山东海洋学院学报,15(1):159-168. Liu Zhongheng, Wu Jinxiu, Yu Yongjie, et al. 1985. The utilization of grain size meterial to preliminary ananlysis of the tidal flay sedimentary environgment of the Huanghe River Delta[J]. Journal of Shandong College of Oceanology, 15(1): 159-168. [5] 孟珊,房景辉,蒋增杰,等. 2020. 双齿围沙蚕对潮间带不同类型底质选择行为的研究[J]. 渔业科学进展,41(4):110-116. Meng Shan, Fang Jinghui, Jiang Zengjie, et al. 2020. Experimental study of the choice behavior of Perinereis aibuhitensis Grube among different sediment types[J]. Progress in Fishery Sciences, 41(4): 110-116. [6] 时硕,吉俊熹,王张华. 2022. 珠江三角洲全新世沉积物C/N和δ 13C变化及对甘蔗种植业的指示[J]. 第四纪研究,42(2):397-411. Shi Shuo, Ji Junxi, Wang Zhanghua. 2022. Holocene variability of bulk organic C/N and δ 13C and implications for the sugarcane cultivation[J]. Quaternary Sciences, 42(2): 397-411. [7] 王翠,王媛媛,胡斌. 2023. 黄河三角洲潮坪环境现代生物遗迹与物化条件的响应关系[J]. 沉积学报,41(3):748-762. Wang Cui, Wang Yuanyuan, Hu Bin. 2023. The response relationship between biogenic structures and physicochemical stresses of the Yellow River deltaic tidal flat[J]. Acta Sedimentologica Sinica, 41(3): 748-762. [8] 王媛媛,勾松林,张国成. 2024. 现代珠江三角洲前缘生物遗迹组成与分布特征[J]. 沉积学报,42(5):1512-1529. Wang Yuanyuan, Gou Songlin, Zhang Guocheng. 2024. Composition and distribution characteristics of biological traces in the Pearl River Delta front[J]. Acta Sedimentologica Sinica, 42(5): 1512-1529. [9] 王媛媛,王学芹,胡斌. 2019. 黄河三角洲潮坪环境中现代生物遗迹组成与分布特征[J]. 沉积学报,37(6):1244-1257. Wang Yuanyuan, Wang Xueqin, Hu Bin. 2019. The composition and distribution characteristics of biogenic sedimentary structures in tidal flat of Yellow River Delta[J]. Acta Sedimentologica Sinica, 37(6): 1244-1257. [10] 韦惺,吴超羽. 2018. 珠江三角洲沉积体与河网干流河道的形成发育[J]. 海洋学报, 40(7):66-78. Wei Xing, Wu Chaoyu. 2018. The formation and development of the deposition bodies and main channels in the Zhujiang River Delta[J]. Acta Oceanologica Sinica, 40(7): 66-78. [11] 曾洪根. 2019. 会爬树的鱼:弹涂鱼[J]. 初中生必读(10):41-42. Zeng Honggen. 2019. Tree climbing fish: Mudskipper[J]. Required Reading for Junior Middle School Students (10): 41-42. [12] 张其永,洪万树,陈仕玺,等. 2006. 潮间带大弹涂鱼产卵习性和自然受精卵孵化研究[J]. 热带海洋学报, 25(5):68-73. Zhang Qiyong, Hong Wanshu, Chen Shixi, et al. 2006. Spawning habits of Boleophthalmus pectinirostris in intertidal mudflat and hatching experiment of natural fertilized eggs[J]. Journal of Tropical Oceanography, 25(5): 68-73. [13] 张绍轩,汤永杰,郑翠美,等. 2020. 珠江三角洲全新世海—陆沉积模式转换及其年代[J]. 海洋地质与第四纪地质,40(5):107-117. Zhang Shaoxuan, Tang Yongjie, Zheng Cuimei, et al. 2020. Holocene sedimentary environment transform and onset time of Pearl River Delta progradation[J]. Marine Geology & Quaternary Geology, 40(5): 107-117. [14] Abdel-Fattah Z A. 2019. Morpho-sedimentary characteristics and generated primary sedimentary structures on the modern microtidal sandy coast of eastern Nile Delta, Egypt[J]. Journal of African Earth Sciences, 150: 355-378. [15] Bann K L, Fielding C R, MacEachern J A, et al. 2004. Differentiation of estuarine and offshore marine deposits using integrated ichnology and sedimentology: Permian Pebbley Beach Formation, Sydney Basin, Australia[J]. Geological Society, London, Special Publications, 228(1): 179-211. [16] Brake S S, Hasiotis S T, Dannelly H K, et al. 2002. Eukaryotic stromatolite builders in acid mine drainage: Implications for Precambrian iron formations and oxygenation of the atmosphere?[J]. Geology, 30(7): 599-602. [17] Carter L, Heath R A. 1975. Role of mean circulation, tides, and waves in the transport of bottom sediment on the New Zealand continental shelf[J]. New Zealand Journal of Marine and Freshwater Research, 9(4): 423-448. [18] Chan F K S, Yang L E, Scheffran J, et al. 2021. Urban flood risks and emerging challenges in a Chinese delta: The case of the Pearl River Delta[J]. Environmental Science & Policy, 122: 101-115. [19] Counts J W, Hasiotis S T. 2009. Neoichnological experiments with Masked Chafer Beetles (Coleoptera: Scarabaeidae): Implications for backfilled continental trace fossils[J]. Palaios, 24(2): 74-91. [20] Dashtgard S E, Gingras M K, Pemberton S G. 2008. Grain-size controls on the occurrence of bioturbation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 257(1/2): 224-243. [21] Drolet D, Bringloe T T, Coffin M R S, et al. 2012. Potential for between-mudflat movement and metapopulation dynamics in an intertidal burrowing amphipod[J]. Marine Ecology Progress Series, 449: 197-209. [22] Geng J J, Wang Y P, Luo H J. 2015. Distribution, sources, and fluxes of heavy metals in the Pearl River Delta, South China[J]. Marine Pollution Bulletin, 101(2): 914-921. [23] Gingras M K, Bann K L, MacEachern J A, et al. 2007. A conceptual framework for the application of trace fossils[M]//MacEachern J A, Bann K L, Gingras M K, et al. Short Course Notes. Tulsa: SEPM: 1-26. [24] Gingras M K, Pemberton S G, Saunders T, et al. 1999. The ichnology of modern and Pleistocene brackish-water deposits at Willapa Bay, Washington: Variability in estuarine settings[J]. Palaios, 14(4): 352-374. [25] Hasiotis S T. 2004. Reconnaissance of Upper Jurassic Morrison Formation ichnofossils, rocky mountain region, USA: Paleoenvironmental, stratigraphic, and paleoclimatic significance of terrestrial and freshwater ichnocoenoses[J]. Sedimentary Geology, 167(3/4): 177-268. [26] Hembree D I, Hasiotis S T. 2006. The identification and interpretation of reptile ichnofossils in paleosols through modern studies[J]. Journal of Sedimentary Research, 76(3): 575-588. [27] Kraus M J, Hasiotis S T. 2006. Significance of different modes of rhizolith preservation to interpreting paleoenvironmental and paleohydrologic settings: Examples from Paleogene paleosols, Bighorn Basin, Wyoming, U.S.A.[J]. Journal of Sedimentary Research, 76(4): 633-646. [28] Kumar A. 2017.Recent biogenic traces from the coastal environments of the southern Red Sea coast of Saudi Arabia[J]. Arabian Journal of Geosciences, 10(22): 500. [29] Lan Z W, Zhang S J, Chen Z Q, et al. 2021.Locomotion and feeding trails produced by crabs[J]. Geological Journal, 56(12): 6288-6293. [30] Lee Y H, Koh C H. 1994. Biogenic sedimentary structures on a Korean mud flat: Spring-neap variations[J]. Netherlands Journal of Sea Research, 32(1): 81-90. [31] Li G, Yan W, Zhong L F. 2016.Element geochemistry of offshore sediments in the northwestern South China Sea and the dispersal of Pearl River sediments[J]. Progress in Oceanography, 141: 17-29. [32] MacEachern J A, Stelck C R, Pemberton S G. 1999. Marine and marginal marine mudstone deposition: Paleoenvironmental interpretations based on the integration of ichnology, palynology and foraminiferal paleoecology[M]//Bergman K M, Snedden J W. Isolated shallow marine sand bodies: Sequence stratigraphic analysis and sedimentologic interpretation. Tulsa: SEPM: 205-225. [33] Mángano M G, Buatois L A. 2004. Ichnology of Carboniferous tide-influenced environments and tidal flat variability in the North American Midcontinent[J]. Geological Society, London, Special Publications, 228(1): 157-178. [34] McIlroy D. 2004. Ichnofabrics and sedimentary facies of a tide-dominated delta: Jurassic Ile Formation of Kristin field, Haltenbanken, offshore Mid-Norway[J]. Geological Society, London, Special Publications, 228(1): 237-272. [35] Miller D J, Eriksson K A. 1997. Late Mississippian prodeltaic rhythmites in the Appalachian Basin; a hierarchical record of tidal and climatic periodicities[J]. Society for Sedimentary Geology, 67(4): 653-660. [36] Nascimento D L, Netto R G, Indicatti R P. 2021. Neoichnology of mygalomorph spiders: Improving the recognition of spider burrows in the geological record[J]. Journal of South American Earth Sciences, 108: 103178. [37] Pemberton S G, Flach P D, Mossop G D. 1982. Trace fossils from the Athabasca oil sands, Alberta, Canada[J]. Science, 217(4562): 825-827. [38] Swinbanks D D, Luternauer J L. 1987. Burrow distribution of thalassinidean shrimp on a Fraser Delta tidal flat, British Columbia[J]. Journal of Paleontology, 61(2): 315-332. [39] Swinbanks D D, Murray J W. 1981. Biosedimentological zonation of Boundary Bay tidal flats, Fraser River Delta, British Columbia[J]. Sedimentology, 28(2): 201-237. [40] Talke S A, Stacey M T. 2003. The influence of oceanic swell on flows over an estuarine intertidal mudflat in San Francisco Bay[J]. Estuarine, Coastal and Shelf Science, 58(3): 541-554. [41] Taylor A, Goldring R, Gowland S. 2003. Analysis and application of ichnofabrics[J]. Earth-Science Reviews, 60(3/4): 227-259. [42] Toniolo M A, Seitz C, Perillo G M E. 2021. Origin and evolution of tidal depressions in a tidal flat and their role in carbon sequestration in the Bahía Blanca Estuary (Argentina)[J]. Marine Geology, 436: 106467. [43] Wang Y Y, Wang X Q, Alfred U, et al. 2019a. Burrows of the polychaete perinereis aibuhiutensis on a tidal flat of the Yellow River Delta in China: Implications for the Ichnofossils Polykladichnus and Archaeonassa[J]. Palaios, 34(5): 271-279. [44] Wang Y Y, Wang X Q, Hu B, et al. 2019b. Tomographic reconstructions of crab burrows from deltaic tidal flat: Contribution to palaeoecology of decapod trace fossils in coastal settings[J]. Palaeoworld, 28(4): 514-524. [45] Weng Q H. 2007. A historical perspective of river basin management in the Pearl River Delta of China[J]. Journal of Environmental Management, 85(4): 1048-1062. [46] Zhang H R, Zhang G F, Jia Q W. 2019. Integration of analytical hierarchy process and landslide susceptibility index based landslide susceptibility assessment of the Pearl River Delta area, China[J]. IEEE Journal of Selected Topics in Applied Earth Observations and Remote Sensing, 12(11): 4239-4251. [47] Zhang S R, Lu X X, Higgitt D L, et al. 2008. Recent changes of water discharge and sediment load in the Zhujiang (Pearl River) Basin, China[J]. Global and Planetary Change, 60(3/4): 365-380. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 856
- HTML全文浏览量: 66
- PDF下载量: 171
- 被引次数: 0

