

 下载:
下载:



-
鲕粒是一种球形或椭球形包覆颗粒(coated grains),由核心和围绕核心的一系列光滑、均匀纹层共同构成。其中,鲕粒核心常见类型包括化石碎片、球粒和陆源碎屑矿物等,纹层主要为钙质成分(CaCO3),少量为磷质、铁质和硅质等。鲕粒粒径一般在0.25~2 mm [1-2]。早期文献常将直径大于2 mm的鲕粒称为豆粒(pisoids),但这种用法遭到Peryt[3]和Richter[2]等人批评。目前国际沉积学界常把粒径大于2 mm的鲕粒称为巨鲕(giant ooids)[4-5],其形貌和内部结构与正常鲕相比并没有明显差别[6]。鲕粒可以形成于不同环境,从海洋、湖泊到洞穴,甚至热泉中均有发现[3,7]。本文主要针对海洋环境下碳酸盐鲕粒展开论述。
目前最古老的鲕粒记录保存在南非的太古宇(约3 000 ~ 2 900 Ma)[8],其出现稍晚于已知最早的生命记录——叠层石(~3 450 Ma)[9]。此后鲕粒广泛发育于碳酸盐主导的沉积环境,并在新元古代、寒武纪、石炭纪、三叠纪,以及侏罗—白垩纪之交分别达到高峰,这与当时表层海水碳酸钙饱和度显著增加有关[10-12]。在现代碳酸盐沉积环境下,鲕粒主要形成于温暖、动荡的浅滩,纬向上主要分布于中—低纬度(大致沿南、北回归线附近)[7,13-15],例如巴哈马[16]和加勒比海[17-18]地区、波斯湾[19]、澳大利亚西海岸(鲨鱼湾)[20],南太平洋社会群岛和库克群岛[21]等地。地质历史时期鲕粒滩的分布与现代类似,但在一些极端气候环境下,也可能出现在整个中、低纬度地区广泛发育的现象(例如早三叠世)[12,22-23]。
鲕粒的净生长速率并不快。以现代巴哈马地区为例,每颗鲕粒每年新增约0.4~18 ng钙质成分,这意味着直径在0.25~0.5 mm之间的鲕粒需要几百到数千年时间才能形成[24-25]。其生长迟缓的原因可能与鲕粒之间碰撞、磨蚀造成的质量损失有关[26]。鲕粒内部结构上表现为同心状、放射状、同心—放射状、泥晶状等多种纹层类型,且不同类型既可以单独发育,也可以共生[27-28]。但是,地质历史时期形成的鲕粒受成岩作用影响并不都能保存下来原始的纹层结构。原生矿物成分的不同是造成鲕粒保存程度存在差异的重要原因之一[27],这主要受控于海水化学性质,表现为以文石和高镁方解石为主(亚稳定矿物)和以低镁方解石为主(稳定矿物)的矿物类型的交替变化,这种变化也被称为“文石海”与“方解石海”之间的转化[27,29]。
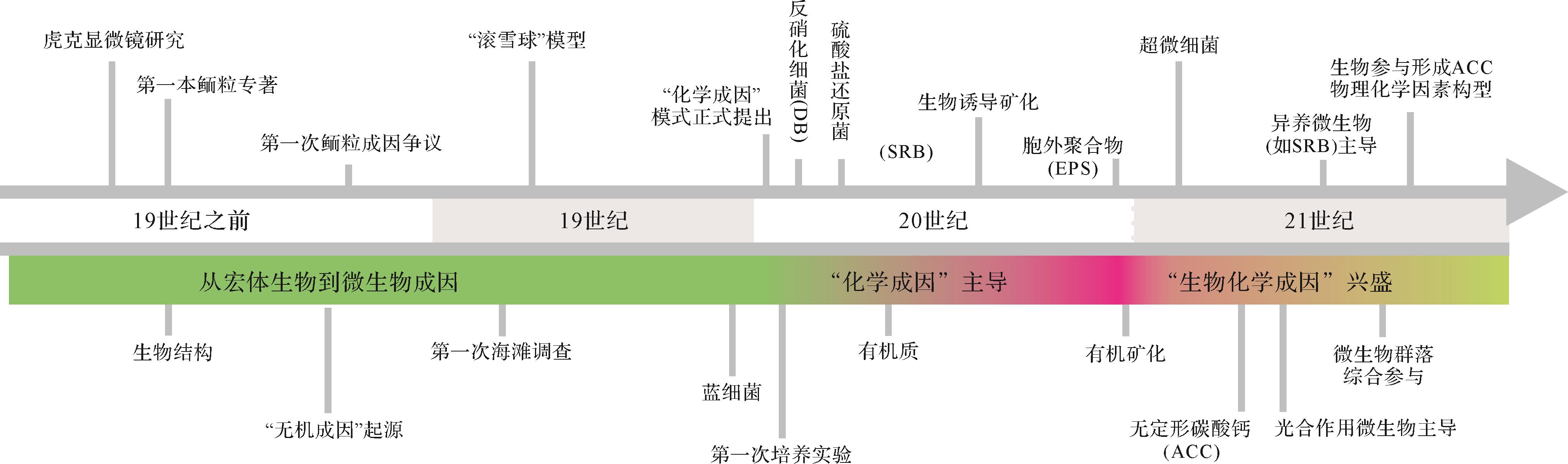
虽然人们对鲕粒在古环境、古气候、古海水性质、储层特征等方面的作用和价值已经有了深入认识,但在鲕粒成因这个最基本问题上,长期以来争议不断(图1)[3,7,10,24,26,29-58]。概括来讲,鲕粒形成主要有物理作用[35],化学作用[38]和生物作用[59]三种主要机制,其中关于生物作用与化学作用哪种机制占主导一直没有定论。目前被大多数学者所接受的“化学作用主导”的认识正受到越来越多生物化学证据的挑战[47-48,51,60]。需要说明的是,关于鲕粒“生物成因”的认识并非是近年来才开始出现的,在鲕粒研究的早期阶段学者们就已经有了“生物参与形成”的朴素认识。随着测试分析技术的不断进步和对有机矿化过程的深入了解[53,61],目前沉积学界对于鲕粒“生物和化学作用共同参与”的认识越来越成熟,大有取代“化学成因”之势(图1)。虽然国外学者对鲕粒成因已经进行过总结[1,3,26,38,43,53,62-65],但这些认识主要是基于对现代鲕粒研究的结果,对于地质历史时期其他类型微生物成因鲕粒阐述有限。国内也有学者对鲕粒成因进行过非常好的总结[66],但相关工作开展较早,对于近十年来鲕粒成因方面的新认识有待梳理。此外,中国有着丰富的鲕粒岩沉积记录,但对于这些鲕粒在成因研究领域的价值还有待深入挖掘。

图 1 鲕粒成因研究主要认识时间线
Figure 1. Timeline of principal insights/views in the study history of the genesis of ooids
基于此,本文从历史和当前视角,回顾了从“藻类参与”到“细菌参与”,以及现在“有机矿化过程”在鲕粒微生物成因研究方面的历程,对鲕粒形成的地球生物学过程进行了系统阐述,并对一些可能与有机矿化过程有关的鲕粒类型进行了探讨,这可为进一步认识鲕粒形成过程、生物与矿物相互作用机理,以及其他具有类似成因的碳酸盐组分研究提供参考。
-
最早对鲕粒进行研究的学者被认为是古罗马博物学家和哲学家Pliny the Elder,他在公元77年的著作Naturalis Historia中首次对鲕粒进行了系统描述,提到鲕粒的形貌与鱼卵有相似之处[34]。之后,Agricola在1546年对鲕粒形态和表面特征进行研究后认为岩石中的鲕粒可类比沙滩上鱼籽的形成与保存[67]。1665年,Hooke[30]在显微镜下首次观察到鲕粒的内部特征,并描述这种结构类似于鸡蛋的同心结构(即,蛋壳、蛋清和蛋黄)。此外,他认为这些颗粒是在流体环境中产生的。Brückmann在1721年出版了关于鲕粒岩的第一本专著[31],进一步提出鲕粒是石化的生物结构[34]。然而,Da Costa在1757年提出,鲕粒起源于微小颗粒的连续结晶,而不是生物化石[32]。1779年,De Saussure[33]也反对“鲕粒来自石化鱼卵”的观点,并进一步指出在动荡水体中细小矿物会不断在鲕粒表面聚集,促进其生长。这两种对立的观点可能是鲕粒成因研究史上生物作用与非生物作用的首次交锋。
自19世纪以来,随着对现代海滩鲕粒的研究,有关其成因的认识得以丰富、完善。De La Beche在1823—1824年对牙买加海滩考察后认为,鲕粒的生长主要依靠碳酸盐过饱和的搅动水环境[68];之后,他提出物理加积和化学沉淀是形成鲕粒的两个重要因素[68]。Dana[69]也持有类似的观点,并进一步强调灰泥的物理加积作用对鲕粒生长的重要性。1879年,著名的鲕粒研究学者Sorby[35]提出鲕粒的生长过程类似于“雪球”的形成,即鲕粒在滚动过程中通过在表面黏附大量细小的颗粒而不断长大。但是,这种观点未被学界广泛接受,因为它无法解释鲕粒光滑的表面结构(很难被黏附)(图2a),以及放射状纹层结构的形成等一系列问题[26,42,70-72]。考虑到鲕粒表面有机物(生物膜)有捕获细小颗粒的能力,一些研究者认为物理加积作用也可作为部分鲕粒形成的影响因素[72-74]。

图 2 现代鲕粒和古代石化鲕粒
Figure 2. Modern ooid sands and ancient fossilized ooids
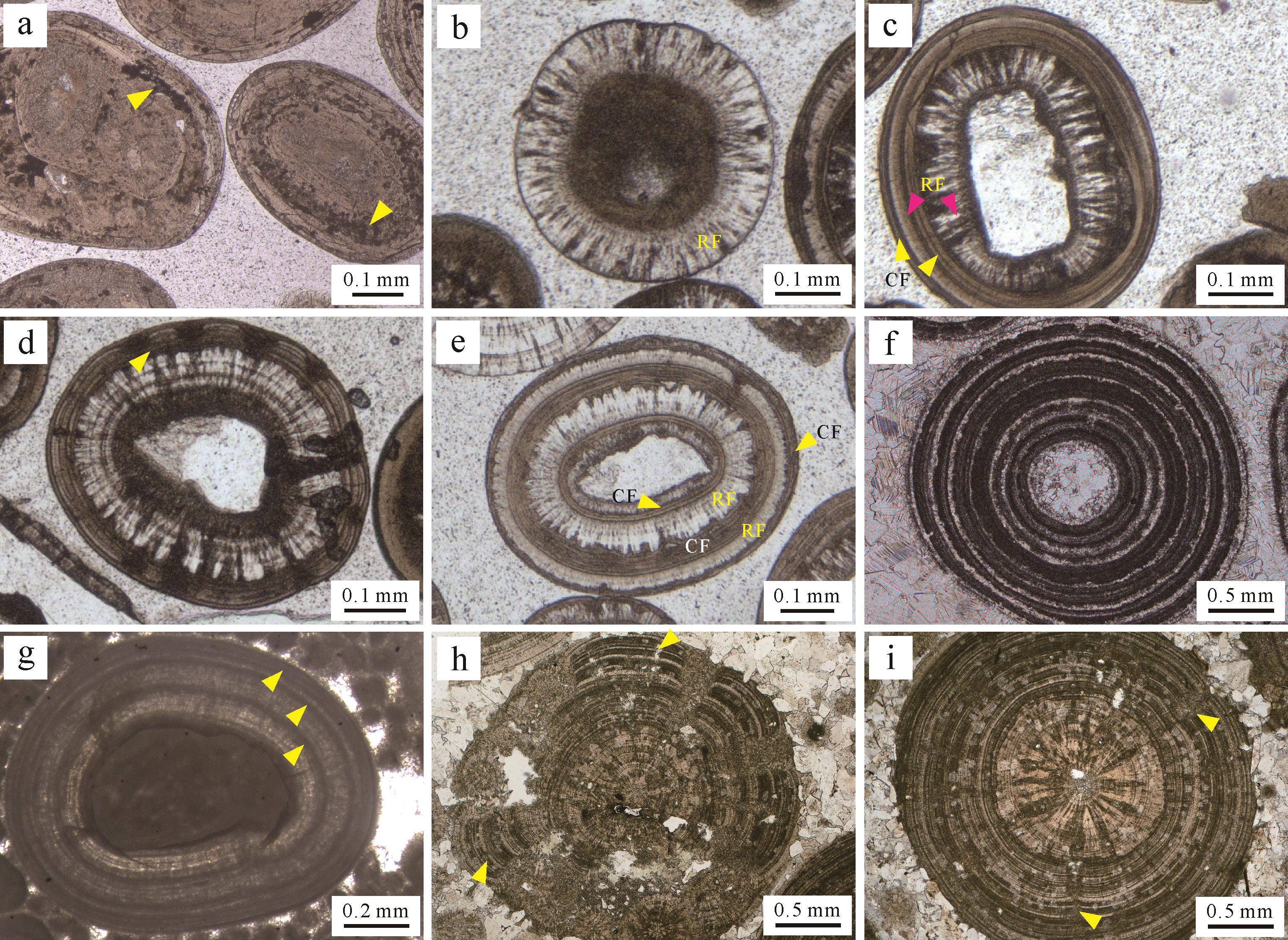
另一方面,鲕粒来源于宏体生物(或其器官、分泌物等)的认识也受到了批评[1,43]。从19世纪末开始,“微藻”在鲕粒形成过程中的价值受到研究者们的关注[36,75]。Wethered[36]和 Rothpletz[37]在研究大盐湖鲕粒时,在与酸性溶液反应后的残渣中识别出一些蓝绿藻(蓝细菌)。Wethered在镜下观察发现这些已经钙化的微生物(如葛万菌等)通常平行或垂直于鲕粒纹层分布,或保存在鲕粒核心[36,75]。基于此,他推测一部分在鲕粒表面生存的蓝细菌具有黏结、捕获周围灰泥的能力,这对于鲕粒生长具有建设性作用;但是其他一些垂直于同心纹层(钻孔类),或在纹层间絮状分布的微生物对鲕粒的形成没有实质性贡献(图3a)。类似的钙化蓝细菌在寒武系鲕粒中也非常常见[76-79]。Kalkowsky[59]在德国哈尔茨山地区(Harz Mountains)下三叠统湖相地层中确认了鲕粒内部存在富有机质纹层,且常具有脑纹状(cerebroid)和条带状(banded)结构,并且这些特征与共生叠层石的纹层结构较为相似(图3h,i)。其中条带状纹层与现代巴哈马鲕粒的同心状纹层之间还存在一定差别:条带状纹层结构通常由暗色和亮色层交替排列,分别对应有机质富集层和贫乏层(图3f)[80];而同心状(切线状)纹层中各个纹层细密且特征相似,不存在间隔变化的特点(图3a)。需要说明的是,地质历史时期鲕粒与叠层石或凝块石的发育既可以相互独立,也可以共存[59,81](图4)。

图 3 典型的鲕粒镜下微结构(单偏光,Leica DM750P显微镜)
Figure 3. Photomicrographs of diagnostic ooid inner microstructures (plane light, Leica DM750P)

图 4 叠层石与鲕粒发育关系
Figure 4. Relationships between microbialite and ooid deposits in sedimentary successions
20世纪初,Linck[38]首次在实验室合成了类似鲕粒的结构(需要注意的是有部分试剂成分也可由生物产生)。其形成过程可以概括为如下反应方程式:
从式中可以看出,鲕粒的生长速率主要取决于水体中Ca2+和
另一方面,随着化学成因模式的兴起,以蓝细菌为代表的“微藻”在鲕粒形成过程中的作用不断受到质疑。由于钻孔活动是蓝细菌的一种常见行为(图3a),这在当时被研究者认为对鲕粒形成主要起破坏作用[26,42,84-86]。此外,在严格无光或不含藻类的环境中,类似鲕粒的结构亦可被合成[87]。考虑到这些不利因素,一些研究者开始探索不进行光合作用的异养型微生物在促进鲕粒纹层形成过程中的可能性[88-89]。需要说明的是,微生物钻孔行为在固定松散碳酸盐颗粒和维护原始纹层结构方面的积极作用在后来得到验证[90]。
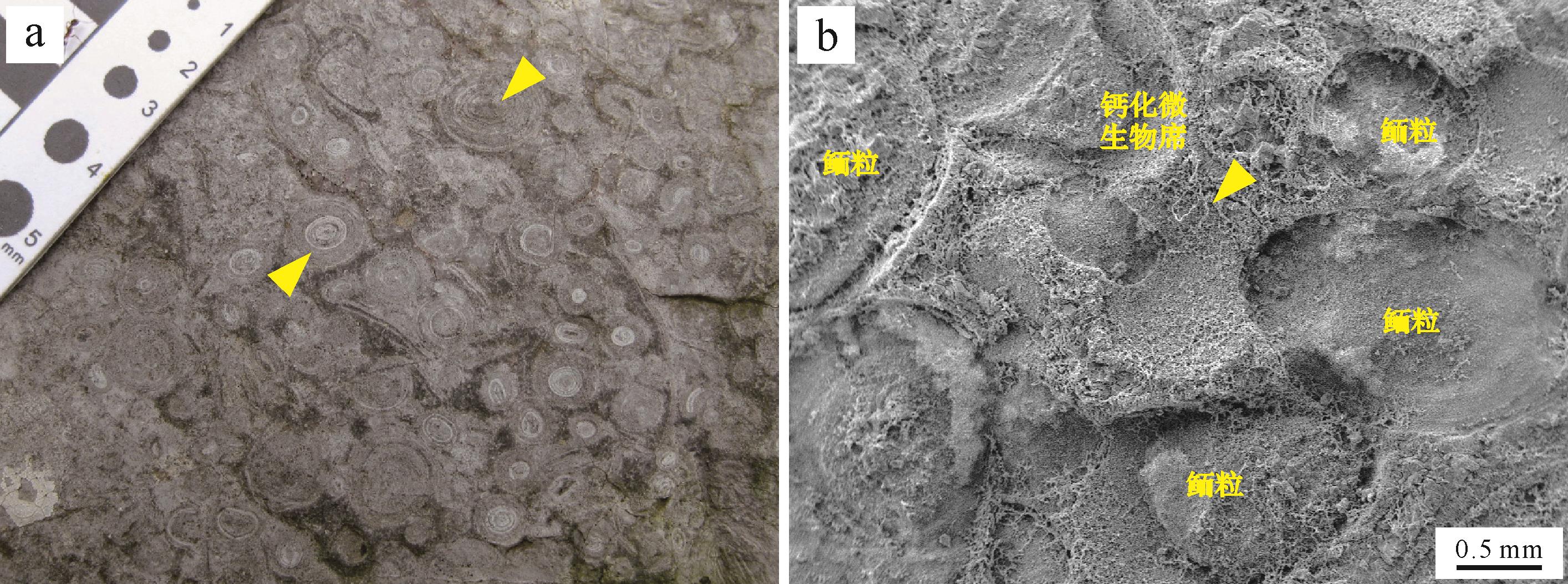
Drew[39,91]在培养实验中引入反硝化细菌后合成了具有类似鲕粒纹层结构的碳酸盐结壳。之后,硫酸盐还原菌在合成类似结构碳酸盐沉积物中的作用也得到了证实[41]。另一方面,一些研究者发现在培养实验中添加一些有机质(如腐殖酸和氨基酸等)[42,86-87]也能形成碳酸钙结晶。进一步的研究确认了有机质和细菌共同参与了碳酸盐的沉淀过程,即一些细菌在代谢过程中通过消耗有机质,在其周围形成碱性微环境诱导碳酸盐成核[26,89,92-94](详见2.2.2节)。Fabricius[43]将这种行为描述为“生物诱导”形成过程,是一种间接的、生物促进矿物形成的机制[95-97],而成核模板(template)主要集中在细菌的胞外聚合物(Extracellular Polymeric Substances,EPS)上。EPS主要为高分子聚合物,在生物诱导矿化过程中主要起到络合和释放金属阳离子,提供有机质来源和成核场所等作用[61]。随后,Gerdes[44]证实了鲕粒纹层内存在大量由细菌和EPS构成的微生物席(膜),这为微生物参与鲕粒形成提供了可供观察的直接证据[98-100]。而鲕粒外部大量发育的微生物席状结构常被作为存在活跃微生物活动以及鲕粒处于停滞期的证据[88](图5)。

图 5 巨鲕和共存的微生物席状结构
Figure 5. Development of giant ooids and associated microbial mats
-
从2000年以来,关于鲕粒成因的研究再次进入一个高潮期,大量新的认识和进展可以概括为如下三个方向。
-
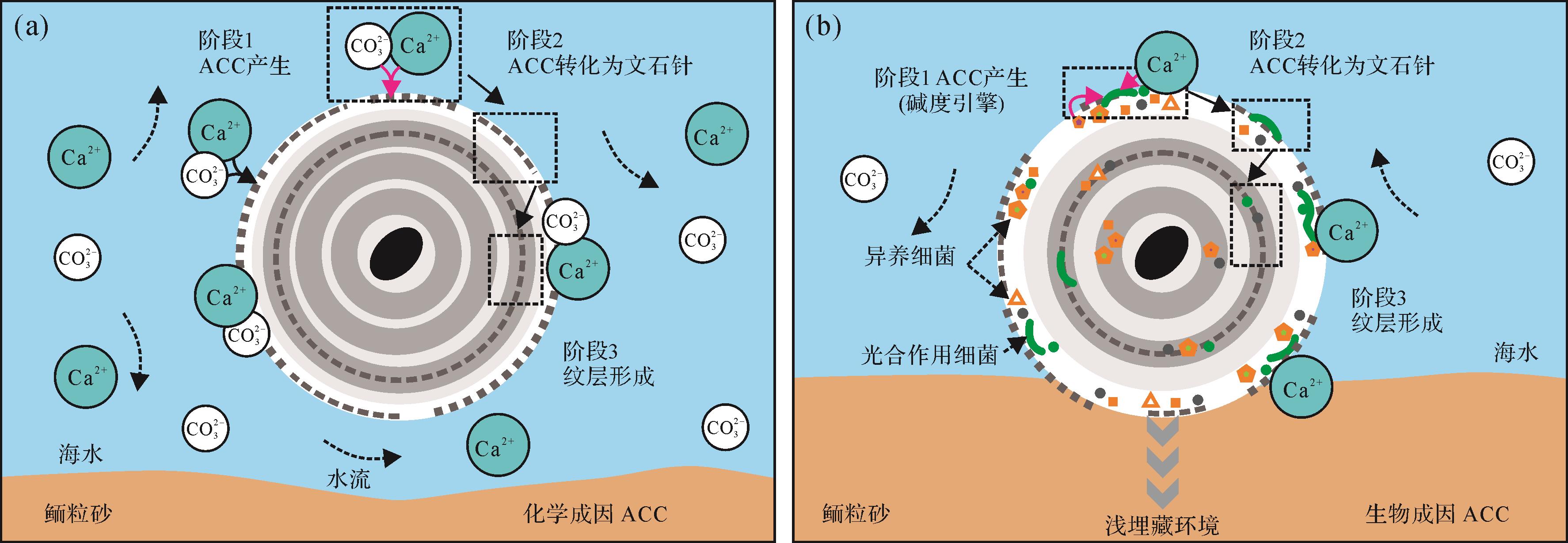
微生物成因矿物的证实为微生物参与鲕粒形成过程提供了直接证据。Folk et al.[46,101-102]通过扫描电子显微镜在鲕粒纹层中识别出大量0.03~0.3 µm的球形或杆状微粒,他们认为这些微粒可能是超微细菌的矿化结构或者是由细菌等产生的纳米级碳酸盐沉淀。实际上,自20世纪70年代以来,这种微小的颗粒就已经被早期鲕粒研究者所关注(如Purser[19]原文图14E;Fabricius[43]原文图11-A2)。但是一些研究人员认为这些超微颗粒的大小(甚至比已知最小的生命体—支原体还要小)和特征不属于微生物的范畴[103-104],倾向于认为这些微粒是纯化学成因或与微生物诱导矿化有关。Duguid et al. [24]认为这些纳米级颗粒(至少一部分)是碳酸盐化学沉淀过程中形成的无定形碳酸钙(ACC),这些ACC随后转化为文石晶体,整个过程不需要微生物参与(图6a)。但是,Diaz et al. [47]通过交叉极化结合魔角旋转核磁共振法技术证实了鲕粒纹层中存在ACC,且这些ACC主要分布于EPS上,暗示了其来源很可能是在有机矿化过程中形成的碳酸钙结晶。随着ACC的大量聚集,在鲕粒外皮层形成薄层。由于ACC不稳定,其随后转变为具有稳定晶形的文石/方解石矿物,但这个过程中晶体排列似乎更多地受到鲕粒周围水化学/物理条件影响(例如,海水Mg/Ca摩尔比值和水动力条件等),进而形成精细纹层结构(图6b)[47,53,105-107]。

图 6 无定形碳酸钙(ACC)在鲕粒纹层形成过程中的两种机制(化学模型和有机矿化模型)
Figure 6. Roles of amorphous calcium carbonates (ACC) in the formation of ooid laminae (Chemical vs. microbial models)
-
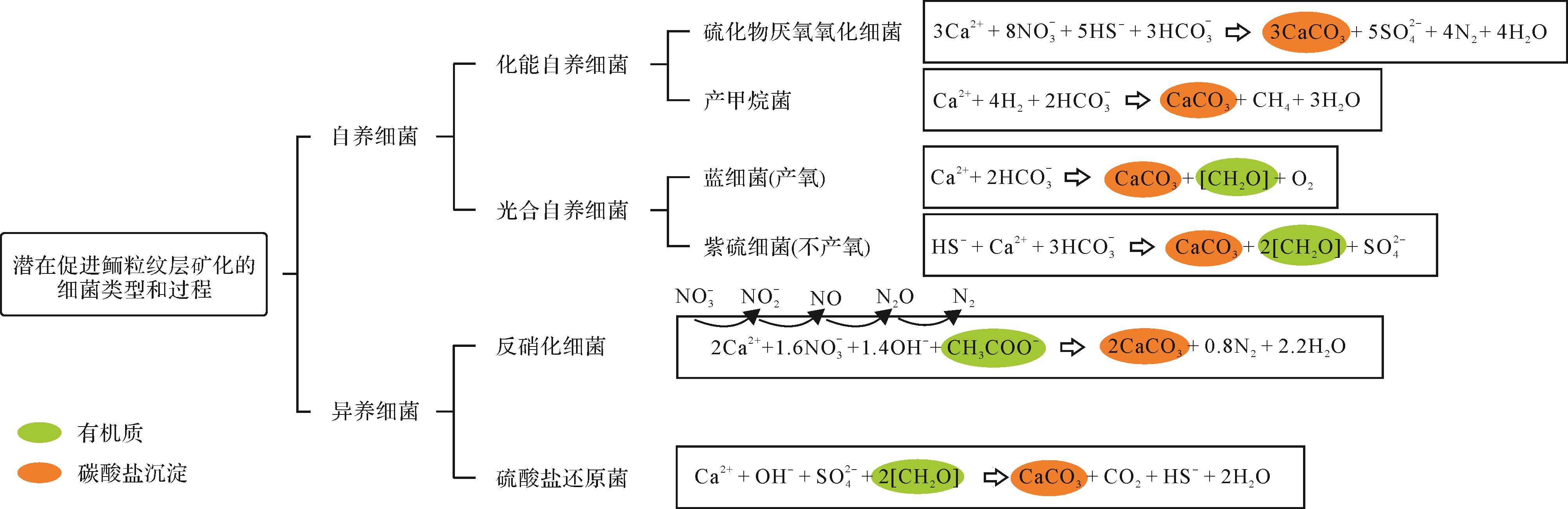
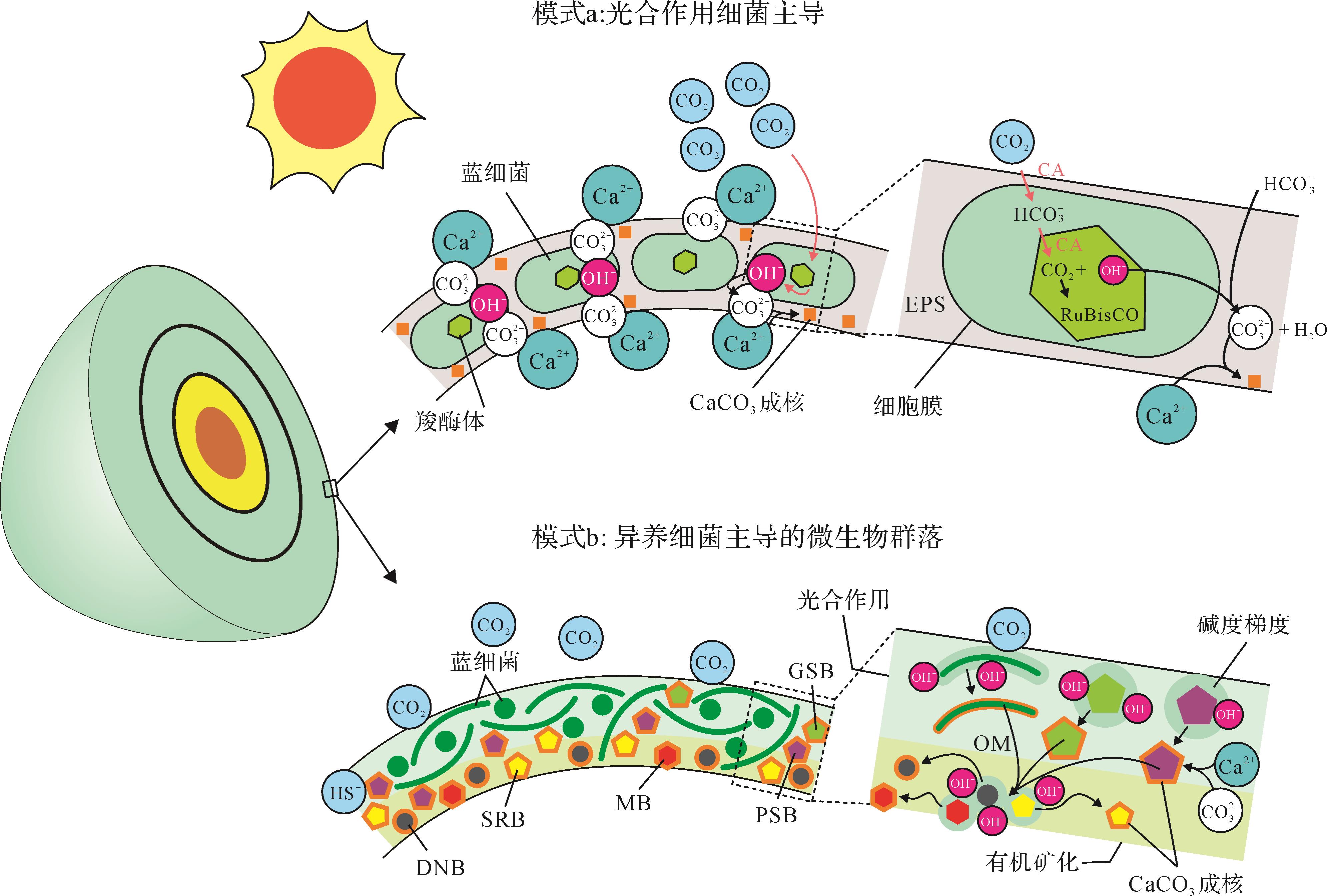
虽然化学沉淀是形成碳酸盐沉积物的一个重要途径,但是有机矿化(生物诱导和生物影响矿化)作用在鲕粒形成过程中同样有可能促进碳酸钙的产生。生物诱导矿化指的是由于生物活动和环境之间相互作用从而产生矿物沉淀的过程[61,108-110],例如蓝细菌光合作用过程中产生的一些副产物(如OH-)造成微环境中碱度或饱和度的增加,进而诱导碳酸盐成核。生物影响矿化指的是由于外界环境条件改变造成碳酸盐在微生物成因沉淀模板(如EPS表面)上成核,与沉淀模板本身无直接关联的一种被动矿化过程[53],例如EPS降解过程中吸附的钙离子释放,造成微环境碳酸钙饱和度增加从而在模板上发生沉淀的过程。目前对于生物诱导矿化作用机理研究较为深入。一些微生物类型,如光合作用细菌(如蓝细菌、紫硫细菌和绿硫细菌等)、硫酸盐还原菌、硫化物厌氧氧化细菌和产甲烷菌等,具有生物诱导矿化能力(图7)。相反,其他能够降低pH和碱度的微生物类型(如硫化物有氧氧化细菌和酵化细菌等)在代谢活动中不利于碳酸盐成核(甚至造成CaCO3损失)[61,111],本文不再论述。就鲕粒形成潜在的贡献者而言,主要包括图7所示类型。

图 7 鲕粒微生物矿化过程中可能存在的微生物类型及其相应的代谢途径(引文详见2.2节)
Figure 7. Types of microbes potentially involved in ooid microbial mineralization and their corresponding metabolic pathways
-
能在鲕粒形成过程中起到促进作用的自养微生物包括化能自养和光能自养细菌两种类型。其中化能自养细菌(如产甲烷菌和硫化物厌氧氧化菌)通常利用H2或还原性硫化物(reduced sulfur compounds)作为电子供体,诱导CaCO3在严格缺氧条件下沉淀[110-111]。研究表明,产甲烷菌和硫化物厌氧氧化菌在各自的代谢过程中每消耗4 mol H2 和5/3 mol HS-便可将1 mol的Ca2+固定下来形成CaCO3沉淀(图7)[111]。光合自养微生物(不产氧和产氧细菌)可将光能转化为化学能进行代谢活动(图7)。其中不产氧光合微生物,如紫硫细菌,利用HS-(由硫酸盐还原作用生成)作为电子供体诱导碳酸盐在氧化还原界面附近成核[109]。其中,紫硫细菌每消耗1 mol HS-便可产生1 mol CaCO3沉淀[111]。产氧光合作用微生物(蓝细菌)在光照下利用H2O(电子供体)固定CO2,并在蓝细菌鞘体(内部或表面)[109]和EPS基质[111-113]上产成CaCO3。蓝细菌在现代湖相鲕粒形成过程中的建造作用是明显的,这些鲕粒纹层中常见到大量保存完好的、钙化的丝状和球状蓝细菌,以及在EPS上形成与生物影响矿化有关的沉淀物[48-50,114]。
尽管如此,蓝细菌作为鲕粒纹层的建造者也存在一些不利因素。首先,蓝细菌的钙化必须同时满足高钙离子(Ca2+)浓度(>13 mmol/L)和低溶解无机碳浓度两个条件[115]。海水中钙离子浓度的降低会极大抑制蓝细菌的钙化,例如,现代海洋中钙离子浓度较低(~10 mmol/L),很难形成钙化蓝细菌[115]。其次,蓝细菌周围EPS似乎不是碳酸盐成核的理想场所,因为在新陈代谢过程中EPS能够吸收周围的Ca2+(和其他离子),从而降低微环境碳酸盐的饱和度[116]。最后,光合作用产生的CaCO3可能被随后的有氧呼吸(好氧异养细菌)耗尽,因而没有产生CaCO3净沉淀[61]。这些因素可能限制了蓝细菌在鲕粒形成中的直接贡献,特别是在现代低钙离子浓度的海洋环境中。
与现代海洋不同的是,湖相微生物岩和鲕粒中常可以保存钙化蓝细菌[48-50,117-118]。其原因可能与:1)周围环境钙离子浓度(或碳酸钙饱和度)较高[112,115];2)富镁硅酸盐基质(镁来自于被分解的EPS(能络合金属离子);硅来自于溶解的硅藻)在蓝细菌细胞/鞘体表面的包覆机制,可抵御周围环境的溶解作用[48,119-121];以及3)协同硫酸盐代谢作用(如不产氧光合作用和硫酸盐还原作用),创造了局部碱性增强的微环境[122]。对于蓝细菌钙化的长期控制因素,Riding[112,123-124]认为CO2浓缩机制可能在大部分地质时期中起着重要作用,该机制的运行可以显著提高CO2吸收率[125],从而诱导鞘层的钙化和EPS的生成[125-126] (图8a)。毫无疑问的是,蓝细菌产生的有机物和附属的EPS是有机质的主要来源之一,可为一些异养微生物(如硫酸盐还原菌和产甲烷菌;图7)新陈代谢活动提供能量来源[61,111,127]。

图 8 鲕粒表皮内微生物矿化的两种模型
Figure 8. Two models of the possible processes of microbial mineralization within ooid cortices
由此可见,蓝细菌在鲕粒纹层形成中的直接作用仍存在争议,但蓝细菌的间接作用,即释放吸附的钙离子和提供有机质来源,对促进其他有机矿化过程起到了重要作用。
-
异养微生物的一些新陈代谢活动同样能够促进碳酸盐沉淀[110],主要包括有机质降解过程中的硝酸盐和硫酸盐的还原[111,128-130]。与氮、硫循环有关的微生物群落被Diaz et al.[52,131]认为在现代巴哈马鲕粒形成过程中起到了主要作用。但是反硝化细菌在微生物诱导碳酸钙沉淀过程中的作用仍存在争议:一些研究人员认为,反硝化作用过程产生CaCO3的效率较高,每消耗1 mol有机质或0.5 mol乙酸盐就可以产生1 mol CaCO3 [132-133];但是也有些研究者认为硝酸盐的还原过程会导致CaCO3的溶解[61,111,134],每消耗1mol有机质将溶解0.2 mol CaCO3。
对于硫酸盐还原菌,大量的研究认为其在诱导碳酸盐沉淀过程中起到了关键作用[111,116,135-139],具体功能包括:1)通过新陈代谢活动营造碱性微环境(pH值增加);2)通过消耗有机质释放已结合的钙离子(主要被EPS吸附);3)通过消除相关的动力学障碍(消耗硫酸盐)促进白云石沉淀。该过程每氧化1 mol有机碳可以产生0.5~1.5 mol的CaCO3沉淀(从不完全氧化到完全氧化)[116]。有意思的是,硫酸盐还原菌并非严格意义上的厌氧微生物,它们也可能出现在有氧环境中[140-141],且某些有氧环境下(如咸化强蒸发)的代谢效率与其在厌氧环境下相当[141-144]。此外,鲕粒生物标志化合物[51]和基因[52,131]分析也证实了硫酸盐还原菌在异养微生物群落中的优势地位。因而一些研究人员认为硫酸盐还原菌可能在鲕粒纹层有机矿化过程中发挥了关键作用(图8b)[51,145]。
-
近年来随着生物标志化合物和基因检测等技术的应用,目前已经证实鲕粒内微生物种群中具有矿化能力的类型与叠层石内非常相似[45,47,52,131,145-148],暗示两者成因可能具有关联。但是目前关于生物矿化过程如何参与鲕粒形成的认识还处于初级阶段。前人认为的微生物参与鲕粒的形成过程主要参考Davies et al. [88]经典的两步生长模型。在这个模型中,受水动力条件和地形影响,鲕粒处于周期性的动荡和停滞状态[16,149-150],当鲕粒脱离强水动力环境并最终被保存在海底时,微生物活动性开始增强(建造或破坏作用)。一些研究者还认为微生物的加积过程可以在特定环境或阶段进行,例如潮下低能环境[150]和鲕粒间歇静止期[114]。在这样的环境中微生物群落可以完成诱导矿化、生物膜(席)建造和初始纹层形成[49-50](图5)。Diaz et al. [47]更是认为生物诱导产生的碳酸钙(ACC形成)可以形成于各个生长阶段,特别是在鲕粒生长活跃期(搅动和间歇静止阶段;图6b)。概括来看,这些观点没有脱离传统的对于鲕粒发育条件的认识,即强水动力和多阶段生长过程控制。
另一类观点不考虑水动力条件的影响,转而强调营养来源和参与鲕粒建造微生物类型的重要性[43-44,151-153]。例如,Brehm et al.[154-155]提出鲕粒是一个具有某些生态优势(例如,利用光照和减少生物捕获)、以蓝细菌为主导的球形微生物群落。这些微生物群落首先构建出一个松散的、具纹层的球状结构,钙质纹层则来源于随后的有机矿化过程。Batchelor et al.[60]进一步推测鲕粒内部微生物群落通过与外界营养交换而不断发育。这些认识在解释具有层状微生物席的现代湖相鲕粒和古代Kalkowsky型鲕粒(类似叠层石构造和富含大量有机质)时似乎更为适合[59-60,156]。但是现代巴哈马鲕粒中很少有类似的层状微生物席的存在(图3a)。
-
鲕粒通常具有排列良好的文石/方解石晶体结构,且化学成因机制可以对CaCO3的沉淀给出令人满意的解释。但是,单一模型并不足以解释在地质历史时期广泛出现的、具有不同内部结构和特征鲕粒的成因。正如许多沉积学家所警告的那样,完全排除微生物在鲕粒形成过程中的建设性作用是不可取的[1,7,34,43,48,51-53,80]。考虑到现阶段关于鲕粒成因的认识仍然存在非常大的争议,研究某些特定类型鲕粒的形成机制可能为解决其成因之谜提供一个突破口。而具有条带状结构的Kalkowsky型鲕粒(图3f,i)则是最有可能存在微生物参与建造的类型。
Kalkowsky型鲕粒在古代海相和湖相地层中非常常见,包括埃迪卡拉系[157-159]、寒武系[78,160]、奥陶系(图3g)、志留系[161]、泥盆系[162]、石炭系[163-165]、三叠系[156,166-170]、侏罗系[170-171]和白垩系[172],且粒径普遍较大(图2b,c、图3f、图5a)。一些现代鲕粒中也具有富含有机质的纹层,如日内瓦湖[48,173-174]、大盐湖[36]和波斯湾[175],其直径一般小于0.5 mm。奇怪的是,巴哈马鲕粒中很少出现有机质富集层。Kalkowsky型鲕粒一般以贫有机质和富有机质纹层的交替出现为特征(图3f,g),其中贫有机质层主要为同心晶粒状和同心—放射纤维状结构(图3c,f)。贫有机质纹层中晶粒状结构很可能是以亚稳定的文石矿物在成岩过程中发生溶解—重结晶改造的结果(图3f)[165,176];而保存条件良好的同心—放射纤维状结构可能与原生矿物为低镁方解石有关(图3g)。此外,脑纹状鲕粒与纹层状鲕粒内部结构高度相似(图3h,i),外凸的纹层结构可能反映了微生物群落对光照和营养的竞争[60]。
Kalkowsky型鲕粒存在有机矿化过程参与基于以下证据:1)富含有机质的纹层由大量钙化的、分层平铺排列且平行于鲕粒表面的微生物残余物(如鞘和EPS)和矿化产物构成(图3f),这些残余物与垂直纹层方向的钻孔微生物特征明显不同;2)富含有机质的纹层相互独立,且与胶结良好的纹层交替出现。这种结构不同于鲕粒在低能环境的泥晶化作用。后者通常会改变鲕粒最外层(或某一层)甚至全部纹层结构(图3a),但很难有规律地间隔改造鲕粒纹层(图3f)。这是由于鲕粒在生长过程中不太可能频繁地在生长阶段和脱离动荡水体的低能环境之间转换;3)贫有机质纹层(颜色亮,晶粒粗)和富有机质纹层(颜色暗,晶粒细)的交替特征与层状叠层石结构高度相似(图2b,c、图3f、图5a)。Burne和Moore称之为“规则纹层状叠层石”(regularly laminational stromatolite),Riding[177]将这种生物作用与化学作用共同参与的叠层石类型称为“混合叠层石”(hybrid stromatolite)。
对于Kalkowsky型鲕粒的形成过程,我们在前人[60,122]的基础上提出了一个改进的模型(图8b)。在这个模型中,鲕粒的表面发育由微生物群落(包括自养和异养微生物)构成的松散微生物席。其中最靠近表层的自养微生物群落(如蓝细菌和绿硫细菌)在光照条件下通过新陈代谢活动不断产生碳酸盐沉淀和有机质,内部的异养微生物(如硫酸盐还原菌和产甲烷菌)通过消耗有机质诱导碳酸盐沉淀。在缺乏光照条件下,以异养微生物群落主导的有机矿化作用将继续参与鲕粒纹层建造。在鲕粒不断生长的过程中,内层的光合自养微生物群落由于光照缺失,在鲕粒纹层形成过程中的直接贡献会消失,从而完全让位给异养微生物主导的有机矿化过程,直到鲕粒表面微生物群落的下一次更新(图6b、图8b)。
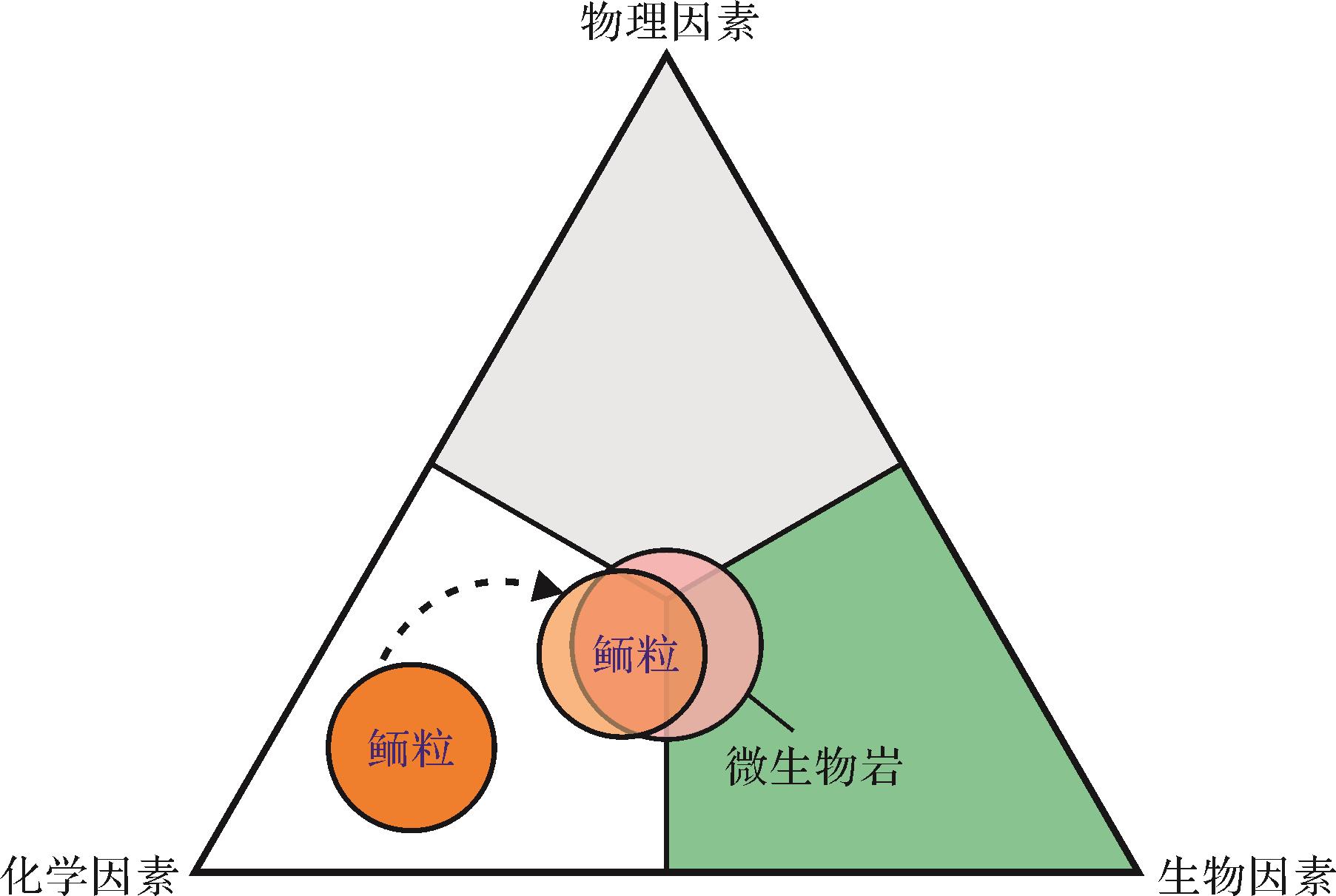
对于Kalkowsky型鲕粒而言,生物过程和化学过程(即生物化学过程)是控制富有机质纹层鲕粒生长的重要因素;相对而言,物理因素的作用是次要的(图9)。而对于同心状和放射状结构的鲕粒,最新的研究认为微生物诱导矿化过程(生物成因ACC的形成)在鲕粒纹层形成的早期阶段非常重要,而从ACC到有序晶体排列的转变可能由物理化学因素控制(图6b)。因此,至少在某些类型(或某些阶段)鲕粒形成过程中,对“完全由化学因素控制鲕粒形成”的传统观念需要再慎重考虑(图9)。

图 9 鲕粒形成过程中的控制因素
Figure 9. Diagram illustrating three interacting factors in the genesis of ooids
-
(1) 从鲕粒成因最新研究认识来看,有机矿化作用产生的ACC以及随后受物理、化学因素影响从ACC到有序晶体排列的转变,可能制约着鲕粒的形成过程。
(2) 考虑到丰度、矿化作用能力和效率等因素,以光合自养细菌(蓝细菌和紫硫细菌)、硫酸盐还原菌和产甲烷菌主导的微生物群落可能在鲕粒有机矿化过程中起到了积极作用。
(3) 不同类型鲕粒中微生物参与的程度可能存在较大差异。Kalkowsky型鲕粒与层状叠层石结构非常类似,其成因可能具有相似性,但有待深入探索。因此,对于与微生物成因有关的鲕粒研究可考虑重点关注富含有机质纹层的Kalkowsky型鲕粒。
Recent Advances in Ooid Microbial Origin: A review
-
摘要:
鲕粒岩在古气候、古环境,以及古海洋领域有着独特的研究价值,然而学术界对于鲕粒成因目前仍没有统一认识。近年来,随着鲕粒形成过程中微生物诱导矿化证据的不断丰富,广为接受的“无机成因”观点正不断受到挑战。本论文从历史和当前视角,回顾了从“藻类参与”到“细菌参与”,以及现在“有机矿化过程”在微生物成因鲕粒研究方面的历程,对鲕粒形成的地球生物学过程进行了系统阐述,并着重介绍了鲕粒形成的各类生物—化学模型。但是这些模型能否解决所有鲕粒的成因问题,还有待进一步工作的检验。考虑到现阶段关于鲕粒成因的认识仍然存在非常大的争议,研究某些特定类型鲕粒的形成机制可为探究鲕粒成因之谜提供一个突破口,而具有条带状纹层结构的Kalkowsky型鲕粒则最有可能存在微生物参与。这是由于Kalkowsky型鲕粒一般以贫有机质和富有机质纹层的交替出现为特征,这与Robert Burne所述的“规则纹层状叠层石”或Robert Riding所述的“混合结构叠层石”特征非常相近,可能意味着它们具有相似的成因。富含有机质的纹层状(或带状)鲕粒虽然在现代巴哈马碳酸盐沉积环境中很少见,但在其他地区和地史记录中较为常见,通过对这种与叠层石特征非常相似的鲕粒进行研究可为了解这类鲕粒纹层形成的有机矿化过程提供了一个新的视角。 Abstract:The study of ooids has a long history, and there is not yet a consensus as to their origin. The “inorganic process” that has been accepted by most sedimentologists has recently been challenged by the accumulating evidence of microbially-mediated mineralization within modern and ancient ooids. For microbial mechanisms involved in the genesis of ooids, photosynthetic microorganisms (e.g., cyanobacteria) use light energy to produce organic carbon and to create alkaline microenvironments during metabolism that induce ambient carbonate nucleation. The organic matter that they produce may feed heterotrophic microorganisms, and some of these heterotrophs (e.g., sulfate-reducing bacteria) are capable of increasing neighboring alkalinity and facilitating carbonate precipitation during their metabolic activities. It is possible that physicochemical conditions rearrange pre-precipitated nanograins produced by biologically-induced and -influenced mineralization processes, and promote the genesis of modern ooids with delicate tangential fabrics (Diaz’s model). In addition, some types of ancient ooids (e.g., alternating dark- and light-colored laminar ooids, and cerebroid ooids) with natural organic matter-rich laminae, closely resemble so-called “regularly laminational stromatolites” or “hybrid stromatolites”, implying their similar origins. The microbial accretion mechanism of such ooids resembles the growth model of laminar stromatolites,i.e., combined biologically-induced (mainly photosynthetic microorganisms and sulfate-reducing bacteria) and biologically-influenced (providing templates for carbonate nucleation) mineralization. Organic matter-rich, laminar (or banded) fabric ooids are rare in modern marine settings but not uncommon in ancient records. Further work focusing on this kind of ooids may provide a new perspective to understand the organomineralization process in the genesis of ooids. -
图 1 鲕粒成因研究主要认识时间线
每个特定发现/观点的引用包括:虎克显微镜研究[30];第一本鲕粒专著[31];生物结构[31];“无机成因”起源[32⁃33];第一次鲕粒成因争议[32⁃33];第一次海滩调查[34];“滚雪球”模型[35];蓝细菌(微藻/蓝绿藻)[36⁃37];“化学成因”模式[38];第一次培养实验[39];反硝化细菌(DB: denitrifying bacteria)参与[40];硫酸盐还原菌(SRB: sulfate⁃reducing bacteria)参与[41];有机质参与[42];生物诱导矿化[43];胞外聚合物(EPS: extracellular polymeric substances)贡献[44];有机矿化作用[45];超微细菌[46];无定形碳酸钙(ACC: amorphous calcium carbonate)[24,47];光合作用微生物主导[48⁃50];异养微生物群落主导[51];微生物群落综合参与[52];生物成因ACC与物理化学因素参与构型[53];具体观点详见第1节;原图参考笔者在《沉积之声》公众号文章《碳酸盐鲕粒工厂》图4
Figure 1. Timeline of principal insights/views in the study history of the genesis of ooids
图 2 现代鲕粒和古代石化鲕粒
(a)巴哈马滩鲕粒(Schooner Cays),具有光滑的外部形貌;(b)寒武系第二统鲕粒(河南龙门石窟);(c)下三叠统鲕粒(广西来宾);其中(b)和(c)均为具有均匀条带状纹层结构的巨鲕
Figure 2. Modern ooid sands and ancient fossilized ooids
图 3 典型的鲕粒镜下微结构(单偏光,Leica DM750P显微镜)
(a)同心状鲕粒。注意钻孔微生物在鲕粒纹层内局部(左侧箭头)和层状(右侧箭头)富集的特征;(b)具放射状(RF: radial fabric)纹层鲕粒;(c)内部放射状(RF)和外部同心状(CF: concentric fabric)结构鲕粒;(d)内部同心放射状和外部脑纹状(轻微,黄色箭头标记)结构鲕粒;(e)放射状(RF)和同心状(CF)结构交替出现的鲕粒;(f)暗色(富有机质)和亮色(贫有机质)纹层交替发育的鲕粒;(g)暗色(富有机质,箭头标记)和亮色(贫有机质,放射—同心状结构)纹层交替的鲕粒;(h)脑纹状结构鲕粒。注意纹层内部含有交替出现的暗色(富有机质)和亮色(贫有机质)纹层(箭头);(i)内部同心—放射状和外部暗色和亮色纹层交替结构。注意出现在外部纹层的轻微脑纹状结构(箭头)。样品来源:(a)现代巴哈马滩;(b~e)现代大盐湖(美国犹他州);(f)广西来宾下三叠统;(g)塔里木盆地上奥陶统;(h~i)德国Harlyberg地区下三叠统
Figure 3. Photomicrographs of diagnostic ooid inner microstructures (plane light, Leica DM750P)
图 4 叠层石与鲕粒发育关系
(a)现代澳大利亚Hamelin Pool叠层石纹层以及纹层之间鲕粒(箭头所指位置)分布特征;(b)河南新乡寒武系苗岭统鲕粒分布于柱状叠层石纹层内部以及叠层石之间(箭头所示);(c)湖北利川下三叠统叠层石纹层(SL: stromatolitic laminae)与纹层间鲕粒层(OL: oolitic laminae),引自文献[81]中图11,经Elsevier授权使用
Figure 4. Relationships between microbialite and ooid deposits in sedimentary successions
图 5 巨鲕和共存的微生物席状结构
(a)下三叠统结构与形貌保存良好的巨鲕(2~8 mm,如箭头所示)。鲕粒一般具有明暗相间的纹层结构;(b)扫描电镜照片显示鲕粒保存环境存在微生物席状结构。注意箭头所示位置保存有大量的EPS,在弱醋酸刻蚀下仍能较好地保存席状结构(鲕粒则呈现凹陷特征);样品来自下三叠统罗楼组(贵州罗甸沫阳)
Figure 5. Development of giant ooids and associated microbial mats
图 6 无定形碳酸钙(ACC)在鲕粒纹层形成过程中的两种机制(化学模型和有机矿化模型)
(a)鲕粒形成的化学模型(参照Davies et al. [88]和Duguid et al. [24]的假设);ACC是由海水沉淀产生,在随后转变为文石针的过程中受到水动力和水化学条件的共同影响;(b)生物成因ACC模型(据Diaz et al. [47,53]);注意生物成因ACC的产生可能发生在动荡的表层海水和/或浅埋藏(暂时停滞)环境中的早期成岩阶段。有机矿化过程中生成的ACC转化为文石针;受水动力和水化学条件的共同影响,鲕粒表层晶体结构发生重新排列
Figure 6. Roles of amorphous calcium carbonates (ACC) in the formation of ooid laminae (Chemical vs. microbial models)
图 7 鲕粒微生物矿化过程中可能存在的微生物类型及其相应的代谢途径(引文详见2.2节)
Figure 7. Types of microbes potentially involved in ooid microbial mineralization and their corresponding metabolic pathways
图 8 鲕粒表皮内微生物矿化的两种模型
(a)CO2浓缩机制调节蓝细菌主导的矿化作用模型[80]。图片经Elsevier授权使用和翻译;(b)异养细菌主导的有机矿化模型。自养细菌与异养细菌共同参与,其中蓝细菌和EPS为有机质重要来源之一[125-126];CA:碳酸酐酶;DNB:反硝化细菌;GSB:绿硫细菌;MB:产甲烷菌;PSB:紫硫细菌;RuBisCO:一种固碳酶;SRB:硫酸盐还原菌。注意可能造成碳酸钙损失的新陈代谢活动(如有氧呼吸、发酵和硫化物有氧氧化)和效率较低的促进碳酸盐沉淀(如氨化)的新陈代谢活动在模型b中没有显示;鲕粒内部的有机矿化过程参考[53,60,108,122]
Figure 8. Two models of the possible processes of microbial mineralization within ooid cortices
-
[1] Simone L. Ooids: A review[J]. Earth-Science Reviews, 1980, 16: 319-355. [2] Richter D K. Calcareous ooids: A synopsis[M]//Peryt T M. Coated grains. Berlin: Springer, 1983: 71-99. [3] Peryt T M. Coated grains[M]. Berlin: Springer, 1983: 1-655. [4] Beukes N J. Ooids and oolites of the proterophytic Boomplaas Formation, Transvaal Supergroup, Griqualand west, South Africa[M]//Peryt T M. Coated grains. Berlin: Springer, 1983: 199-214. [5] Sumner D Y, Grotzinger J P. Numerical modeling of ooid size and the problem of Neoproterozoic giant ooids[J]. Journal of Sedimentary Research, 1993, 63(5): 974-982. [6] 李飞,王夏,薛武强,等. 一种新的错时相沉积物:巨鲕及其环境意义[J]. 沉积学报,2010,28(3):585-595. Li Fei, Wang Xia, Xue Wuqiang, et al. Origin and environmental significance of giant ooids in the Early Triassic: A new kind of anachronistic facies[J]. Acta Sedimentologica Sinica, 2010, 28(3): 585-595. [7] Tucker M E, Wright V P. Carbonate sedimentology[M]. Oxford: Blackwell Science, 1990: 1-496. [8] Siahi M, Hofmann A, Master S, et al. Carbonate ooids of the Mesoarchaean Pongola Supergroup, South Africa[J]. Geobiology, 2017, 15(6): 750-766. [9] Hofmann H J, Grey K, Hickman A H, et al. Origin of 3.45 Ga coniform stromatolites in Warrawoona Group, western Australia[J]. GSA Bulletin, 1999, 111(8): 1256-1262. [10] Wilkinson B H, Owen R M, Carroll A R. Submarine hydrothermal weathering, global eustasy, and carbonate polymorphism in Phanerozoic marine oolites[J]. Journal of Sedimentary Research, 1985, 55(2): 171-183. [11] Li F, Yan J X, Chen Z Q, et al. Global oolite deposits across the Permian-Triassic boundary: A synthesis and implications for palaeoceanography immediately after the end-Permian biocrisis[J]. Earth-Science Reviews, 2015, 149: 163-180. [12] Opdyke B N, Wilkinson B H. Paleolatitude distribution of Phanerozoic marine ooids and cements[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 1990, 78(1/2): 135-148. [13] Lees A, Buller A T. Modern temperate-water and warm-water shelf carbonate sediments contrasted[J]. Marine Geology, 1972, 13(5): M67-M73. [14] Lippmann F. Sedimentary carbonate minerals[M]. Berlin: Springer, 1973: 1-229. [15] Gallagher S J, Reuning L, Himmler T, et al. The enigma of rare Quaternary oolites in the Indian and Pacific Oceans: A result of global oceanographic physicochemical conditions or a sampling bias?[J]. Quaternary Science Reviews, 2018, 200: 114-122. [16] Reeder S L, Rankey E C. Interactions between tidal flows and ooid shoals, northern Bahamas[J]. Journal of Sedimentary Research, 2008, 78(3): 175-186. [17] Ward W C, Brady M J. High-energy carbonates on the inner shelf, northeastern Yucatan Peninsula, Mexico[J]. GCAGS Transactions, 1973, 23: 226-238. [18] Trower E J, Cantine M D, Gomes M L, et al. Active ooid growth driven by sediment transport in a high-energy shoal, Little Ambergris Cay, Turks and Caicos Islands[J]. Journal of Sedimentary Research, 2018, 88(9): 1132-1151. [19] Purser B H. The Persian Gulf: Holocene carbonate sedimentation and diagenesis in a shallow Epicontinental sea[M]. Berlin: Springer, 1973: 1-474. [20] Logan B W, Davies G R, Read J F, et al. Carbonate sedimentation and environments, Shark Bay, western Australia[M]. Tulsa: American Association of Petroleum Geologists, 1970: 1-205. [21] Rankey E C, Reeder S L. Holocene ooids of Aitutaki Atoll, Cook Islands, South Pacific[J]. Geology, 2009, 37(11): 971-974. [22] Witzke B J. Palaeoclimatic constraints for Palaeozoic palaeolatitudes of Laurentia and Euramerica[M]//McKerrow W S, Scotese C R. Palaeozoic palaeogeography and biogeography. London: Geological Society, 1990: 57-73. [23] Li F, Gong Q L, Burne R V, et al. Ooid factories operating under hothouse conditions in the earliest Triassic of South China[J]. Global and Planetary Change, 2019, 172: 336-354. [24] Duguid S M A, Kyser T K, James N P, et al. Microbes and ooids[J]. Journal of Sedimentary Research, 2010, 80(3): 236-251. [25] Beaupré S R, Roberts M L, Burton J R, et al. Rapid, high-resolution 14C chronology of ooids[J]. Geochimica et Cosmochimica Acta, 2015, 159: 126-138. [26] Bathurst R G C. Carbonate sediments and their diagenesis[M]. Amsterdam: Elsevier Science, 1972: 1-658. [27] 李飞,武思琴,刘柯. 鲕粒原生矿物识别及对海水化学成分变化的指示意义[J]. 沉积学报,2015,33(3):500-511. Li Fei, Wu Siqin, Liu Ke. Identification of ooid primary mineralogy: A clue for understanding the variation in paleo-oceanic chemistry[J]. Acta Sedimentologica Sinica, 2015, 33(3): 500-511. [28] 宋文天,刘建波. 碳酸盐鲕粒包壳结构研究综述[J]. 古地理学报,2020,22(1):147-160. Song Wentian, Liu Jianbo. A review of cortical structures of carbonate ooids[J]. Journal of Palaeogeography, 2020, 22(1): 147-160. [29] Sandberg P A. An oscillating trend in Phanerozoic non-skeletal carbonate mineralogy[J]. Nature, 1983, 305(5929): 19-22. [30] Hooke R. Micrographia, or, Some physiological descriptions of minute bodies made by magnifying glasses, with observations and inquiries thereupon[M]. London: J. Martyn and J. Allestry, 1665: 1-246. [31] Brückmann F E. Specimen physicum exhibens historiam naturalem, oolithi seu ovariorum piscium & concharum in saxa mutatorum[M]. Helmestadii: Salomoni & Schnorrii, 1721: 1-28. [32] Da Costa E M. A natural history of fossils[M]. London: L. Davis and C. Reymers, 1757: 1-294. [33] De Saussure H B. Voyages dans les Alpes, précédés d'un essai sur l'histoire naturelle des environs de Genève[M]. Neuchâtel: Franche-Borel, 1779: 1-540. [34] Burne R V, Eade J C, Paul J. The natural history of ooliths: Franz Ernst Brückmann’s treatise of 1721 and its significance for the understanding of oolites[J]. Hallesches Jahrbuch für Geowissenschaften, 2012, 34: 93-114. [35] Sorby H C. The structure and origin of limestones[J]. Proceedings of the Geological Society of London, 1879, 35: 56-95. [36] Wethered E. On the occurrence of the genus Girvanella in oolitic rocks, and remarks in oolitic structure[J]. Quarterly Journal of the Geological Society, 1890, 46(1/2/3/4): 270-283. [37] Rothpletz A. On the formation of oolite[J]. Botanisches Centralblatt, 1892, 10(5): 279-283. [38] Linck G. Die bildung der oolithe und rogensteine[J]. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 1903, 16: 495-513. [39] Drew G H. The action of some denitrifying bacteria in tropical and temperate seas, and the bacterial precipitation of calcium carbonate in the sea[J]. Journal of the Marine Biological Association of the United Kingdom, 1911, 9(2): 142-155. [40] Vaughan T W. Preliminary remarks on the geology of the Bahamas, with special reference to the origin of the Bahaman and Floridian oolites[J]. Washington: Tortugas Laboratory of the Carnegie Institution of Washington, 1914, 102: 47-54. [41] Monaghan P H, Lytle M L. The origin of calcareous ooliths[J]. Journal of Sedimentary Research, 1956, 26(2): 111-118. [42] Newell N D, Purdy E G, Imbrie J. Bahamian oölitic sand[J]. The Journal of Geology, 1960, 68(5): 481-497. [43] Fabricius F H. Origin of marine oöids and grapestones[M]. Stuttgart: E. Schweizerbart'sche Verlagsbuchhandlung, 1977: 1-113. [44] Gerdes G, Dunajtschik-Piewak K, Riege H, et al. Structural diversity of biogenic carbonate particles in microbial mats[J]. Sedimentology, 1994, 41(6): 1273-1294. [45] Reitner J, Arp G, Thiel V, et al. Organic matter in Great Salt Lake ooids (Utah, USA) - First approach to a formation via organic matrices[J]. Facies, 1997, 36: 210-219. [46] Folk R L. SEM imaging of bacteria and nannobacteria in carbonate sediments and rocks[J]. Journal of Sedimentary Research, 1993, 63(5): 990-999. [47] Diaz M R, Eberli G P, Blackwelder P, et al. Microbially mediated organomineralization in the formation of ooids[J]. Geology, 2017, 45(9): 771-774. [48] Pacton M, Ariztegui D, Wacey D, et al. Going nano: A new step toward understanding the processes governing freshwater ooid formation[J]. Geology, 2012, 40(6): 547-550. [49] Plee K, Ariztegui D, Martini R, et al. Unravelling the microbial role in ooid formation - results of an in situ experiment in modern freshwater Lake Geneva in Switzerland[J]. Geobiology, 2008, 6(4): 341-350. [50] Plée K, Pacton M, Ariztegui D. Discriminating the role of photosynthetic and heterotrophic microbes triggering low-Mg calcite precipitation in freshwater biofilms (Lake Geneva, Switzerland)[J]. Geomicrobiology Journal, 2010, 27(5): 391-399. [51] Summons R E, Bird L R, Gillespie A L, et al. Lipid biomarkers in ooids from different locations and ages: Evidence for a common bacterial flora[J]. Geobiology, 2013, 11(5): 420-436. [52] Diaz M R, Van Norstrand J D, Eberli G P, et al. Functional gene diversity of oolitic sands from Great Bahama Bank[J]. Geobiology, 2014, 12(3): 231-249. [53] Diaz M R, Eberli G P. Decoding the mechanism of formation in marine ooids: A review[J]. Earth-Science Reviews, 2019, 190: 536-556. [54] Budd D A, Land L S. Geochemical imprint of meteoric diagenesis in Holocene ooid sands, Schooner Cays, Bahamas: Correlation of calcite cement geochemistry with extant groundwaters[J]. Journal of Sedimentary Research, 1990, 60(3): 361-378. [55] Heydari E, Snelling R D, Dawson W C, et al. Ooid mineralogy and diagenesis of the Pitkin Formation, North-central Arkansas[M]//Keith B D, Zuppann C W. Mississippian oolites and modern analogs. Tulsa: American Association of Petroleum Geologists, 1993: 175-184. [56] Kidder D L, Hall S. Petrology and diagenetic evolution of Neoproterozoic ooids (Libby Formation, western Montana, U.S.A.)[J]. Precambrian Research, 1993, 63(1/2): 83-96. [57] Harris P M. Facies anatomy and diagenesis of a Bahamian ooid shoal[M]. Miami: University of Miami, 1979: 1-150. [58] Li F, Webb G E, Algeo T J, et al. Modern carbonate ooids preserve ambient aqueous REE signatures[J]. Chemical Geology, 2019, 509: 163-177. [59] Kalkowsky E. Oolith und Stromatolith im norddeutschen Buntsandstein[J]. Zeitschrift der Deutschen Geologischen Gesellschaft Band, 1908, 60: 68-125. [60] Batchelor M T, Burne R V, Henry B I, et al. A biofilm and organomineralisation model for the growth and limiting size of ooids[J]. Scientific Reports, 2018, 8(1): 559. [61] Dupraz C, Reid R P, Braissant O, et al. Processes of carbonate precipitation in modern microbial mats[J]. Earth-Science Reviews, 2009, 96(3): 141-162. [62] Brown T C. Origin of oolites and the oolitic texture in rocks[J]. GSA Bulletin, 1914, 25(1): 745-780. [63] Illing L V. Bahaman calcareous sands[J]. AAPG Bulletin, 1954, 38(1): 1-95. [64] Eardley A J. Sediments of Great Salt Lake, Utah[J]. AAPG Bulletin, 1938, 22(10): 1305-1411. [65] Harris P, Diaz M R, Eberli G P. The formation and distribution of modern ooids on Great Bahama Bank[J]. Annual Review of Marine Science, 2019, 11: 491-516. [66] 梅冥相. 鲕粒成因研究的新进展[J]. 沉积学报,2012,30(1):20-32. Mei Mingxiang. Brief introduction on new advances on the origin of ooids[J]. Acta Sedimentologica Sinica, 2012, 30(1): 20-32. [67] Bandy M C, Bandy J A. De natura fossilium (textbook of mineralogy)[M]. New York: The Geological Society, 1955: 1-240. [68] De La Beche H T. The geological observer[M]. Philadelphia: Blanchard and Lea, 1851: 1-695. [69] Dana J D. Corals and coral islands[M]. New York: Dodd Mead & Co, 1890: 1-398. [70] Bathurst R G C. Precipitation of oöids and other aragonite fabrics in warm seas[M]//Müller G, Friedman G M. Recent developments in carbonate sedimentology in Central Europe. Berlin: Springer, 1968: 1-10. [71] Land L S, Behrens E W, Frishman S A. The ooids of Baffin Bay, Texas[J]. Journal of Sedimentary Research, 1979, 49(4): 1269-1277. [72] Morse J W, Mackenzie F T. Geochemistry of sedimentary carbonates[M]. Amsterdam: Elsevier, 1990: 1-725. [73] Shearman D J, Twyman J, Karimi M Z. The genesis and diagenesis of oolites[J]. Proceedings of the Geologists' Association, 1970, 81(3): 561-564, IN7-IN9, 565-575. [74] Folk R L. Carbonate petrography in the post-Sorbian age[M]//Ginsburg R N. Evolving concepts in sedimentology. Baltimore: Johns Hopkins University Press, 1973: 118-158. [75] Wethered E B. The formation of oolite[J]. Quarterly Journal of the Geological Society, 1895, 51(1/2/3/4): 196-209. [76] Spincer B R. Oolitized fragments of filamentous calcimicrobes and the pseudofossil affinity of Nuia Maslov from the Upper Cambrian rocks of central Texas[J]. Journal of Paleontology, 1998, 72(3): 577-584. [77] Liu W, Zhang X L. Girvanella-coated grains from Cambrian oolitic limestone[J]. Facies, 2012, 58(4): 779-787. [78] 代明月,齐永安,陈尧,等. 豫西渑池地区寒武系第三统张夏组的巨鲕及其成因[J]. 古地理学报,2014,16(5):726-734. Dai Mingyue, Qi Yong’an, Chen Yao, et al. Giant ooids and their genetic analysis from the Zhangxia Formation of Cambrian Series 3 in Mianchi area, western Henan province[J]. Journal of Palaeogeography, 2014, 16(5): 726-734. [79] Han Z Z, Zhang X L, Chi N J, et al. Cambrian oncoids and other microbial-related grains on the North China Platform[J]. Carbonates and Evaporites, 2015, 30(4): 373-386. [80] Li F, Yan J X, Burne R V, et al. Paleo-seawater REE compositions and microbial signatures preserved in laminae of Lower Triassic ooids[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 486: 96-107. [81] Fang Y H, Chen Z Q, Kershaw S, et al. An Early Triassic (Smithian) stromatolite associated with giant ooid banks from Lichuan (Hubei province), South China: Environment and controls on its formation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2017, 486: 108-122. [82] Cayeux L. Les roches sédimentaires de France: Roches carbonatées[M]. Paris: Masson, 1935: 1-463. [83] Sandberg P A. New interpretations of Great Salt Lake ooids and of ancient non-skeletal carbonate mineralogy[J]. Sedimentology, 1975, 22(4): 497-537. [84] Klement K W, Toomey D F. Role of the blue-green alga Girvanella in skeletal grain destruction and lime-mud formation in the Lower Ordovician of West Texas[J]. Journal of Sedimentary Research, 1967, 37(4): 1045-1051. [85] Golubic S, Seong-Joo L, Browne K M. Cyanobacteria: Architects of sedimentary structures[M]//Riding R E, Awramik S M. Microbial sediments. Berlin: Springer, 2000: 57-67. [86] Shearman D J, P A d'E Skipwith. Organic matter in recent and ancient limestones and its role in their diagenesis[J]. Nature, 1965, 208(5017): 1310-1311. [87] Mitterer R M. Amino acid composition of organic matrix in calcareous oolites[J]. Science, 1968, 162(3861): 1498-1499. [88] Davies P J, Bubela B, Ferguson J. The formation of ooids[J]. Sedimentology, 1978, 25(5): 703-730. [89] Ferguson J, Bubela B, Davies P J. Synthesis and possible mechanism of formation of radial carbonate ooids[J]. Chemical Geology, 1978, 22: 285-308. [90] Reid R P, Macintyre I G. Microboring versus recrystallization: Further insight into the micritization process[J]. Journal of Sedimentary Research, 2000, 70(1): 24-28. [91] Drew G H. On the precipitation of calcium carbonate in the sea by marine bacteria, and on the action of denitrifying bacteria in tropical and temperate seas[J]. Journal of the Marine Biological Association of the United Kingdom, 1913, 9(4): 479-524. [92] Mitterer R M. Biogeochemistry of aragonite mad and oolites[J]. Geochimica et Cosmochimica Acta, 1972, 36(12): 1407-1422. [93] Suess E, Futterer D. Aragonitic ooids: Experimental precipitation from seawater in the presence of humic acid[J]. Sedimentology, 1972, 19(1/2): 129-139. [94] Trichet J. Etude de la composition de la fraction organique des oolites. Comparaison avec celle des membranes des bactéries et des cyanophycées[J]. Comptes rendus de l'Académie des Sciences de Paris, Série D, 1968, 267: 1492-1494. [95] Krumbein W E. Photolithotropic and chemoorganotrophic activity of bacteria and algae as related to beachrock formation and degradation (gulf of Aqaba, Sinai)[J]. Geomicrobiology Journal, 1979, 1(2): 139-203. [96] Krumbein W E. Calcification by bacteria and algae[M]//Trudinger P A, Swaine D J. Biogeochemical cycling of mineral-forming elements. Amsterdam: Elsevier, 1979: 47-68. [97] Krumbein W E, Cohen Y, Shilo M. Solar Lake (Sinai). 4. Stromatolitic cyanobacterial mats[J]. Limnology and Oceanography, 1977, 22(4): 635-656. [98] Decho A W. Microbial exopolymer secretions in ocean environments: Their role(s) in food webs and marine processes[J]. Oceanography and Marine Biology: An Annual Review, 1990, 28: 73-153. [99] Decho A W, Visscher P T, Reid R P. Production and cycling of natural microbial exopolymers (EPS) within a marine stromatolite[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 219(1/2): 71-86. [100] Gallagher K L, Dupraz C, Braissant O, et al. Mineralization of sedimentary biofilms: Modern mechanistic insights[M]//Bailey W C. Biofilms: Formation, development and properties. New York: Nova Science Publishers, 2010: 227-258. [101] Folk R L, Leo Lynch F. Organic matter, putative nannobacteria and the formation of ooids and hardgrounds[J]. Sedimentology, 2001, 48(2): 215-229. [102] Folk R L, Chafetz H S. Bacterially induced microscale and nanoscale carbonate precipitates[M]//Riding R E, Awramik S M. Microbial sediments. Berlin: Springer, 2000: 40-49. [103] Young J D, Martel J. The rise and fall of nanobacteria[J]. Scientific American, 2010, 302(1): 52-59. [104] Southam G. A structural comparison of bacterial microfossils vs. 'nanobacteria' and nanofossils[J]. Earth-Science Reviews, 1999, 48(4): 251-264. [105] Obst M, Dynes J J, Lawrence J R, et al. Precipitation of amorphous CaCO3 (aragonite-like) by cyanobacteria: A STXM study of the influence of EPS on the nucleation process[J]. Geochimica et Cosmochimica Acta, 2009, 73(14): 4180-4198. [106] Benzerara K, Menguy N, López-García P, et al. Nanoscale detection of organic signatures in carbonate microbialites[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(25): 9440-9445. [107] Couradeau E, Benzerara K, Gérard E, et al. An early-branching microbialite cyanobacterium forms intracellular carbonates[J]. Science, 2012, 336(6080): 459-462. [108] Diaz M R, Swart P K, Eberli G P, et al. Geochemical evidence of microbial activity within ooids[J]. Sedimentology, 2015, 62(7): 2090-2112. [109] Konhauser K, Riding R. Bacterial biomineralization[M]//Knoll A H, Canfield D E, Konhauser K O. Fundamentals of geobiology. Chichester: John Wiley & Sons, 2012: 105-130. [110] Castanier S, Le Métayer-Levrel G, Perthuisot J P. Bacterial roles in the precipitation of carbonate minerals[M]//Riding R E, Awramik S M. Microbial sediments. Berlin: Springer, 2000: 32-39. [111] Visscher P T, Stolz J F. Microbial mats as bioreactors: Populations, processes, and products[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 219(1/2): 87-100. [112] Riding R. Calcified cyanobacteria[M]//Reitner J, Thiel V. Encyclopedia of geobiology. Dordrecht: Springer, 2011: 211-223. [113] Riding R. Temporal variation in calcification in marine cyanobacteria[J]. Journal of the Geological Society, 1992, 149(6): 979-989. [114] Davaud E, Girardclos S. Recent freshwater ooids and oncoids from western Lake Geneva (Switzerland): Indications of a common organically mediated origin[J]. Journal of Sedimentary Research, 2001, 71(3): 423-429. [115] Arp G, Reimer A, Reitner J. Photosynthesis-induced biofilm calcification and calcium concentrations in Phanerozoic oceans[J]. Science, 2001, 292(5522): 1701-1704. [116] Dupraz C, Visscher P T. Microbial lithification in marine stromatolites and hypersaline mats[J]. Trends in Microbiology, 2005, 13(9): 429-438. [117] Arp G, Reimer A, Reitner J. Microbialite formation in seawater of increased alkalinity, Satonda Crater Lake, Indonesia[J]. Journal of Sedimentary Research, 2003, 73(1): 105-127. [118] Couradeau E, Benzerara K, Gérard E, et al. Cyanobacterial calcification in modern microbialites at the submicrometer scale[J]. Biogeosciences, 2013, 10(8): 5255-5266. [119] Wacey D, Urosevic L, Saunders M, et al. Mineralisation of filamentous cyanobacteria in Lake Thetis stromatolites, western Australia[J]. Geobiology, 2018, 16(2): 203-215. [120] Burne R V, Moore L S, Christy A G, et al. Stevensite in the modern thrombolites of Lake Clifton, western Australia: A missing link in microbialite mineralization?[J]. Geology, 2014, 42(7): 575-578. [121] Souza-Egipsy V, Wierzchos J, Ascaso C, et al. Mg–silica precipitation in fossilization mechanisms of sand tufa endolithic microbial community, Mono Lake (California)[J]. Chemical Geology, 2005, 217(1/2): 77-87. [122] Pace A, Bourillot R, Bouton A, et al. Formation of stromatolite lamina at the interface of oxygenic-anoxygenic photosynthesis[J]. Geobiology, 2018, 16(4): 378-398. [123] Riding R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic–Cambrian changes in atmospheric composition[J]. Geobiology, 2006, 4(4): 299-316. [124] Riding R. An atmospheric stimulus for cyanobacterial-bioinduced calcification ca. 350 million years ago?[J]. PALAIOS, 2009, 24(10): 685-696. [125] Huisman J, Codd G A, Paerl H W, et al. Cyanobacterial blooms[J]. Nature Reviews Microbiology, 2018, 16(8): 471-483. [126] Raven J A, Beardall J, Sánchez-Baracaldo P. The possible evolution and future of CO2-concentrating mechanisms[J]. Journal of Experimental Botany, 2017, 68(14): 3701-3716. [127] Tang D J, Shi X Y, Shi Q, et al. Organomineralization in Mesoproterozoic giant ooids[J]. Journal of Asian Earth Sciences, 2015, 107: 195-211. [128] Soetaert K, Hofmann A F, Middelburg J J, et al. Reprint of “The effect of biogeochemical processes on pH”[J]. Marine Chemistry, 2007, 106(1/2): 380-401. [129] Canfield D E, Thamdrup B. Towards a consistent classification scheme for geochemical environments, or, why we wish the term 'suboxic' would go away[J]. Geobiology, 2009, 7(4): 385-392. [130] Dupraz, C, Visscher, P T, Baumgartner, L K, Reid, R P. Microbe–mineral interactions: early carbonate precipitation in a hypersaline lake (Eleuthera Island, Bahamas)[J]. Sedimentology, 2004, 51(4): 745-765. [131] Diaz M R, Piggot A M, Eberli G P, et al. Bacterial community of oolitic carbonate sediments of the Bahamas Archipelago[J]. Marine Ecology Progress Series, 2013, 485: 9-24. [132] Erşan Y Ç, De Belie N, Boon N. Microbially induced CaCO3 precipitation through denitrification: An optimization study in minimal nutrient environment[J]. Biochemical Engineering Journal, 2015, 101: 108-118. [133] Hamdan N, Kavazanjian Jr E, Rittmann B E, et al. Carbonate mineral precipitation for soil improvement through microbial denitrification[J]. Geomicrobiology Journal, 2017, 34(2): 139-146. [134] Rassmann J, Lansard B, Pozzato L, et al. Carbonate chemistry in sediment porewaters of the Rhône River delta driven by early diagenesis (northwestern Mediterranean)[J]. Biogeosciences, 2016, 13(18): 5379-5394. [135] Canfield D E, Des Marais D J. Biogeochemical cycles of carbon, sulfur, and free oxygen in a microbial mat[J]. Geochimica et Cosmochimica Acta, 1993, 57(16): 3971-3984. [136] Lyons W B, Long D T, Hines M E, et al. Calcification of cyanobacterial mats in Solar Lake, Sinai[J]. Geology, 1984, 12(10): 623-626. [137] Castanier S, Le Métayer-Levrel G, Perthuisot J P. Ca-carbonates precipitation and limestone genesis — the microbiogeologist point of view[J]. Sedimentary Geology, 1999, 126(1/2/3/4): 9-23. [138] Braissant O, Decho A W, Dupraz C, et al. Exopolymeric substances of sulfate-reducing bacteria: Interactions with calcium at alkaline pH and implication for formation of carbonate minerals[J]. Geobiology, 2007, 5(4): 401-411. [139] Wright D T. The role of sulphate-reducing bacteria and cyanobacteria in dolomite formation in distal ephemeral lakes of the Coorong region, South Australia[J]. Sedimentary Geology, 1999, 126(1/2/3/4): 147-157. [140] Canfield D E, Des Marais D J. Aerobic sulfate reduction in microbial mats[J]. Science, 1991, 251(5000): 1471-1473. [141] Fründ C, Cohen Y. Diurnal cycles of sulfate reduction under oxic conditions in cyanobacterial mats[J]. Applied and Environmental Microbiology, 1992, 58(1): 70-77. [142] Baumgartner L K, Reid R P, Dupraz C, et al. Sulfate reducing bacteria in microbial mats: Changing paradigms, new discoveries[J]. Sedimentary Geology, 2006, 185(3/4): 131-145. [143] Teske A, Ramsing N B, Habicht K, et al. Sulfate-reducing bacteria and their activities in cyanobacterial mats of Solar Lake (Sinai, Egypt)[J]. Applied and Environmental Microbiology, 1998, 64(8): 2943-2951. [144] Caumette P, Matheron R, Raymond N, et al. Microbial mats in the hypersaline ponds of Mediterranean salterns (Salins-de-Giraud, France)[J]. FEMS Microbiology Ecology, 1994, 13(4): 273-286. [145] O'reilly S S, Mariotti G, Winter A R, et al. Molecular biosignatures reveal common benthic microbial sources of organic matter in ooids and grapestones from Pigeon Cay, The Bahamas[J]. Geobiology, 2017, 15(1): 112-130. [146] Revsbech N P, Jorgensen B B, Blackburn T H, et al. Microelectrode studies of the photosynthesis and O2, H2S, and pH profiles of a microbial mat[J]. Limnology and Oceanography, 1983, 28(6): 1062-1074. [147] Visscher P T, Prins R A, Van Gemerden H. Rates of sulfate reduction and thiosulfate consumption in a marine microbial mat[J]. FEMS Microbiology Letters, 1992, 86(4): 283-293. [148] Edgcomb V P, Bernhard J M, Beaudoin D, et al. Molecular indicators of microbial diversity in oolitic sands of Highborne Cay, Bahamas[J]. Geobiology, 2013, 11(3): 234-251. [149] Anderson N T, Cowan C A, Bergmann K D. A case for the growth of ancient ooids within the sediment pile[J]. Journal of Sedimentary Research, 2020, 90: 843-854. [150] Mariotti G, Pruss S B, Summons R E, et al. Contribution of benthic processes to the growth of ooids on a low-energy shore in Cat Island, The Bahamas[J]. Minerals, 2018, 8(6): 252. [151] Dahanayake K, Gerdes G, Krumbein W E. Stromatolites, oncolites and oolites biogenically formed in situ[J]. Naturwissenschaften, 1985, 72(10): 513-518. [152] Kahle C F J. Proposed origin of aragonite Bahaman and some Pleistocene marine ooids involving bacteria, nannobacteria(?), and biofilms[J]. Carbonates and Evaporites, 2007, 22(1): 10-22. [153] Jones B, Goodbody Q H. Biological factors in the formation of quiet-water ooids[J]. Bulletin of Canadian Petroleum Geology, 1984, 32(2): 190-200. [154] Brehm U, Krumbein W E, Palinska K A. Biomicrospheres generate ooids in the laboratory[J]. Geomicrobiology Journal, 2006, 23(7): 545-550. [155] Brehm U, Palinska K A, Krumbein W E. Laboratory cultures of calcifying biomicrospheres generate ooids - A contribution to the origin of oolites[J]. Carnets de Géologie, 2004, CG2004(L03): 1-6. [156] Paul J, Peryt T M. Kalkowsky's stromatolites revisited (Lower Triassic Buntsandstein, Harz Mountains, Germany)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2000, 161(3/4): 435-458. [157] Thorie A, Mukhopadhyay A, Banerjee T, et al. Giant ooids in a Neoproterozoic carbonate shelf, Simla Group, Lesser Himalaya, India: An analogue related to Neoproterozoic glacial deposits[J]. Marine and Petroleum Geology, 2018, 98: 582-606. [158] Corsetti F A, Kidder D L, Marenco P J. Trends in oolite dolomitization across the Neoproterozoic–Cambrian boundary: A case study from Death Valley, California[J]. Sedimentary Geology, 2006, 191(3/4): 135-150. [159] Lu C J, Li F, Oehlert A M, et al. Reconstructing paleoceanographic conditions during the middle Ediacaran: Evidence from giant ooids in South China[J]. Precambrian Research, 2020, 351: 105945. [160] Tan Q, Shi Z J, Tian Y M, et al. Origin of ooids in ooidal-muddy laminites: A case study of the Lower Cambrian Qingxudong Formation in the Sichuan Basin, South China[J]. Geological Journal, 2018, 53(5): 1716-1727. [161] Stel J H, De Coo J C. The Silurian Upper burgsvik and Lower Hamra-Sundre beds, Gotland[J]. Scripta Geologica, 1977, 44: 1-43. [162] Playton T E, Kerans C. Late Devonian carbonate margins and Foreslopes of the Lennard shelf, Canning Basin, western Australia, part A: Development during backstepping and the aggradation-to-progradation transition[J]. Journal of Sedimentary Research, 2015, 85(11): 1334-1361. [163] Webb G E. Quantitative analysis and paleoecology of earliest Mississippian microbial reefs, Gudman Formation, Queensland, Australia: Not just post-disaster phenomena[J]. Journal of Sedimentary Research, 2005, 75(5): 877-896. [164] Pollard A. Origin of Visean (Mississippian) Frobisher Group coated grainstone shoals in the Williston Basin: Deposition and diagenesis[D]. Kingston: Queen’s University, 2017: 1-81. [165] Algeo T J, Watson B A. Calcite, aragonite, and bimineralic ooids in Missourian (Upper Pennsylvanian) strata of Kansas: Stratigraphic and geographic patterns of variation[M]//Pausé P H, Candelaria M P. Carbonate facies and sequence stratigraphy: Practical applications of carbonate models. Tulsa: PBGC-SEPM Publication, 1995: 141-173. [166] Li F, Yan J X, Algeo T, et al. Paleoceanographic conditions following the End-Permian mass extinction recorded by giant ooids (Moyang, South China)[J]. Global and Planetary Change, 2013, 105: 102-120. [167] Gu Y F, Jiang Y Q, Lei X H, et al. The major controlling factors and different oolitic shoal reservoir characteristics of the Triassic Feixianguan Formation, eastern Longgang area, NE Sichuan Basin, SW China[J]. Acta Geologica Sinica (English Edition), 2021, 95(3): 895-908. [168] Dozet S, Ogorelec B. Younger Paleozoic, Mesozoic and Tertiary oolitic and oncolitic beds in Slovenia – an overview[J]. Geologija, 2012, 55(2): 181-208. [169] Tian L, Bottjer D J, Tong J N, et al. Distribution and size variation of ooids in the aftermath of the Permian-Triassic mass extinction[J]. Palaios, 2015, 30(9): 714-727. [170] Ogorelec B. Mikrofacies mezozojskih karbonathnih kamnin Slovenije[M]. Ljubljana: Geološki zavod Slovenije, 2011: 1-136. [171] Reolid M, Gaillard C, Lathuilière B. Microfacies, microtaphonomic traits and foraminiferal assemblages from Upper Jurassic oolitic–coral limestones: Stratigraphic fluctuations in a shallowing-upward sequence (French Jura, Middle Oxfordian)[J]. Facies, 2007, 53(4): 553-574. [172] Suarez-Gonzalez P, Quijada I E, Benito M I, et al. Do stromatolites need tides to trap ooids? Insights from a Cretaceous system of coastal-wetlands[M]//Tessier B, Reynaud J Y. Contributions to modern and ancient tidal sedimentology: Proceedings of the tidalites 2012 conference. Chichester: John Wiley & Sons, 2016: 161-190. [173] Ariztegui D, Plee K, Farah R, et al. Bridging the gap between biological and sedimentological processes in ooid formation: Crystalizing F.A. FOREL's vision[J]. Archives des Sciences, 2012, 65: 93-102. [174] Plee K. Geomicrobiological investigation of ooid cortex formation in a freshwater environment-Lake Geneva, Switzerland[D]. Genève: Université de Genève, 2008: 1-137. [175] Loreau J P, Purser B H. Distribution and ultrastructure of Holocene ooids in the Persian Gulf[M]//Purser B H. The Persian gulf: Holocene carbonate sedimentation and diagenesis in a Shallow Epicontinental Sea. Berlin: Springer, 1973: 279-328. [176] Assereto R, Folk R L. Brick-like texture and radial rays in Triassic pisolites of Lombardy, Italy: A clue to distinguish ancient aragonitic pisolites[J]. Sedimentary Geology, 1976, 16(3): 205-222. [177] Riding R. Microbialites, stromatolites, and thrombolites[M]//Reitner J, Thiel V. Encyclopedia of geobiology. Dordrecht: Springer, 2011: 635-654. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1132
- HTML全文浏览量: 354
- PDF下载量: 444
- 被引次数: 0

