

 下载:
下载:

-
埃迪卡拉纪到寒武纪是重要的地质历史时期,其间发生了构造、海洋、生物等一系列共同变化[1⁃2]。其中,以早寒武世骨骼动物爆发性出现,即“寒武纪生命大爆发”最引人关注。目前关于这一事件的触发机制仍不明确。通常认为这一时期海洋的整体氧化为生物爆发创造了必要条件[3⁃5]。然而大量基于铁组分、氧化还原敏感微量元素等地球化学指标的研究却显示了华南在这一时期海洋依然以强烈的氧化还原分层为主[6⁃7]。基于目前对华南氮同位素统计,Wang et al.[8]认为埃迪卡拉生物群、小壳生物群和澄江生物群的出现均对应于硝酸盐浓度的提升阶段,而埃迪卡拉生物群及小壳生物群灭绝则与硝酸盐浓度下降有关,据此提出埃迪卡拉—早寒武世生命演化可能受控于硝酸盐浓度的动态变化。然而,Chang et al.[9]对含有清江生物群(大致与澄江生物群等时)地层的古环境重建结果显示这一时期古海洋依然具有很低的硝酸盐浓度,因此质疑了硝酸盐浓度在寒武纪生命大爆发中的作用。此外,虽然晚埃迪卡拉纪古海洋具有较高的硝酸盐浓度,但同期也仅有埃迪卡拉生物群,并未出现大规模的生物爆发现象[10⁃11]。因此,查明早寒武世古海洋氧化还原结构及营养物水平,对深入认识早寒武世生物大爆发具有重要意义。
作为重要的生命元素,氮组分(硝酸盐/铵根)的多寡及相对比例对生命演化/演替具有重要影响[12⁃13],其循环过程又与古海洋氧化还原状态/结构密切相关。因此,对重大地史时期开展氮同位素研究能同时获悉生产力变化及古海洋氧化还原条件两方面信息。此外,由于氮同位素可以反映化变层的相对位置[14],若和反映底水氧化还原状态的指标(如铁组分,微量元素等)相结合,能够更好地刻画古海洋的氧化还原结构及演化。目前,前人对华南早寒武世氮同位素已开展部分研究工作[9,15⁃18]。虽然这些研究总体显示了华南早寒武世较低的氮同位素特征,但同时揭示了该时期氮循环的强烈时空非均质性。一方面,华南早寒武世的陆架区和斜坡—盆地区显示了相反的氮同位素演化模式(例如,陆架区正偏,斜坡—盆地区负偏);另一方面,华南早寒武世氮循环具有固氮、反硝化、铵同化等多种组合模式。因此,有必要采用多指标联合的方法对这一时期的氮循环进行进一步的约束。
以扬子东南缘秀山地区ZK0202井为研究对象,系统采集下寒武统牛蹄塘组及变马冲组底部的岩心样品,通过主量元素、微量元素、稀土元素、碳—氮同位素等指标分析,重建了研究区早寒武世古海洋氧化还原环境,在此基础上进一步探讨了同期氮循环模式,以期更好地厘清早寒武世生物大爆发的古海洋环境因素与形成机制。
-
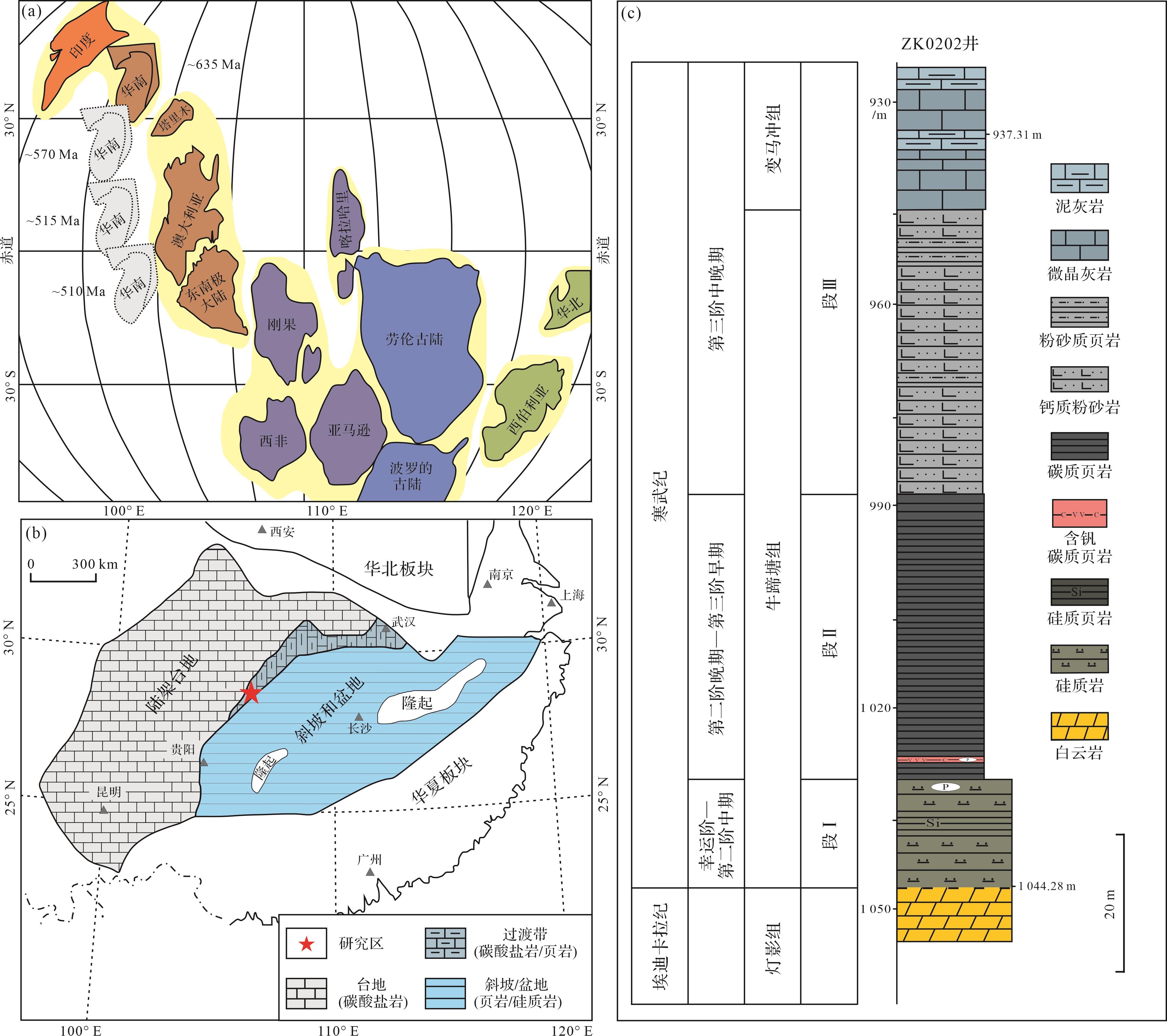
早寒武世,华南克拉通位于赤道附近(图1a),主要由华夏板块和扬子板块组成[21]。受新元古代构造事件的影响,华南克拉通由裂谷盆地逐渐向被动大陆边缘演化[22]。华南继承了新元古代的古地理格局,从北西向南东方向依次发育浅水陆架台地相、台地边缘相、斜坡—盆地相(图1b)。台地相主要发育白云岩、灰岩[24],台地边缘相以页岩、灰岩互层为主[25],而斜坡—盆地相则多发育硅质岩和页岩[26⁃27]。

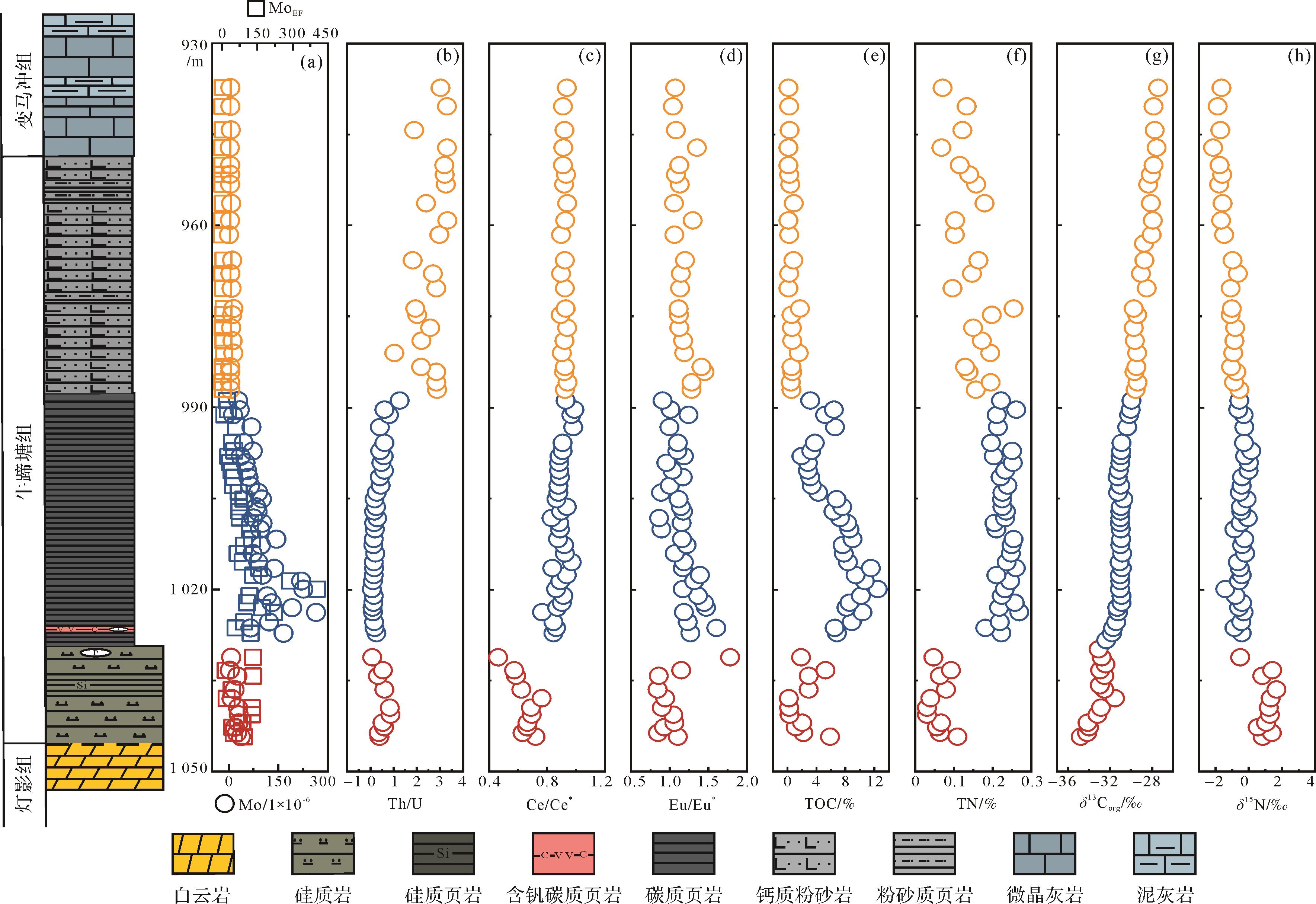
图 1 (a)寒武纪早期全球古地理图(据文献[19⁃20]修改);(b)寒武纪早期扬子台地古地理图(据文献[23]修改);(c)ZK0202井牛蹄塘组—变马冲组岩性柱
Figure 1. (a) Early Cambrian global paleogeography (modified from references [19⁃20]); (b) paleogeographic map of the Yangtze Platform during the early Cambrian (modified from reference [23]; (c) lithological column of the Niutitang Formationand Bianmachong Formation in well ZK0202)
重点解剖研究钻孔(ZK0202井)位于扬子板块东南缘渝东南秀山地区,古地理位置属于上扬子板块的陆架—斜坡过渡带。岩心自下而上发育埃迪卡拉纪陡山沱组、灯影组;早寒武世牛蹄塘组、变马冲组、金顶山组、清虚洞组。其中,牛蹄塘组总体上可分为底部的硅质岩段和中上部泥岩段(图1c)。底部硅质岩厚15.28 m,偶夹硅质页岩,顶部含磷结核,与下伏灯影组平行不整合接触;中上部泥页岩段可进一步分为下部黑色页岩段(40.52 m)和上部粉砂质泥岩段(39.58 m)。黑色页岩段底部出现含钒(V)金属矿层,其余以碳质页岩为主;粉砂质泥岩段向上进一步出现微晶灰岩,属于变马冲组(图1c)。
根据岩性特征和前人的研究成果[24,28],将岩心分为三段。其中段Ⅰ为硅质岩段,基于生物地层学、年代地层学和岩石地层学的证据,上扬子地区的黑色页岩首现层位(SSF3出现)为526 Ma[29⁃31],因此将ZK0202井黑色页岩底部限定为526 Ma,则硅质岩段均早于526 Ma,属于寒武纪幸运阶—第二阶中期(>526 Ma);段Ⅱ为黑色页岩段,其中V矿层出现年龄大致为521 Ma,常被认为与Ni-Mo矿层等时[8,28,32],这进一步支持了黑色页岩底部年龄的限定。另外,基于生物地层(三叶虫和澄江生物群)和岩石地层学证据,黑色页岩向粉砂岩转变的层位被大致限定为518 Ma[33⁃34]。同时,这一层位也出现了总有机碳(TOC)含量的显著下降。因此,将ZK0202井黑色页岩向粉砂岩转变层位的年龄大致限定为518 Ma,属于寒武纪第二阶晚期—寒武纪第三阶早期(ca. 526~518 Ma);段Ⅲ为粉砂岩至微晶灰岩,属于寒武纪第三阶中晚期(<518 Ma)。
-
研究样品均来自ZK0202井的新鲜岩心样品,共计71个样品,采样深度为937.31~1 044.28 m。所有样品进行了有机碳同位素(δ13Corg)测试。挑选出66个样品进行总有机碳(TOC)、总氮(TN)、主微量元素、稀土元素含量及氮同位素(δ15N)测试。所有样品均小心避开方解石脉,切成小块之后研磨成均匀的粉末,以备后续分析。样品的TOC含量、TN含量、δ13Corg测试均在中国科学院广州地球化学研究所有机地球化学国家重点实验室完成。δ15N测试在自然资源部第三海洋研究所海洋生物与生态实验室分析完成。主量及微量元素测试在武汉上谱分析科技有限责任公司完成。
-
称取约0.1 g样品粉末置于陶瓷坩埚中,加入1∶1盐酸去除碳酸盐组分,随后用去离子水清洗至中性,并在60 ℃下烘干。TOC含量利用LECO CS-344碳硫分析仪进行分析。使用实验室内部标准B4016(TC=2.24%)进行数据检测,分析精度优于0.1%。根据TOC含量,称取适量的去碳酸盐样品,进行δ13Corg测试。测试仪器为MAT253同位素比质谱仪,使用实验室内部标样CBW04408(δ13Corg=-36.91‰)进行数据检测,分析精度优于0.2‰。
-
称取约0.1 g样品粉末放入离心管中,加入1∶1盐酸去除碳酸盐组分。称取约15 mg已除去碳酸盐的样品进行TN(TNdecarb)含量测试。测试仪器为VARIO EL cube元素分析仪,使用实验室内部标样IVA3380(TC=1.86%,TN=0.122%)进行数据检测,分析精度优于0.3%。根据记录的处理前后样品质量差,计算原始样品中TN含量。
根据TNdecarb含量称取10~15 mg去碳酸盐样品,进行δ15N分析。测试仪器为Thermo Fisher Delta V Plus气体同位素比质谱仪,使用标样ACET(δ15N=-4.21‰)进行数据检测,分析精度优于0.3‰。
-
全岩主量元素的分析方法为熔片法。将全岩样品放入马弗炉中煅烧去除有机质,记录烧矢量;然后称取0.5 g煅烧后样品以质量比1∶8与Li2B4O7均匀混合,利用Rigaku ZSX Primus Ⅱ型波长色散X射线荧光光谱仪(XRF)进行分析,各主量元素分析谱线均为Kα,标准曲线使用国家标准物质岩石系列:GBW07101-14、土壤系列:GSS07401-08、水系沉积物系列:GBW07302-12建立。数据校正采用理论α系数法,测试相对标准偏差(RSD)小于2%。全岩微量元素含量利用Agilent 7700e ICP-MS分析完成。样品处理如下:(1)将200目样品置于105 ℃烘箱中烘干12 h;(2)准确称取粉末样品50 mg置于Teflon溶样弹中;(3)先后依次缓慢加入1 mL高纯HNO3和1 mL高纯HF;(4)将Teflon溶样弹放入钢套,拧紧后置于190 ℃烘箱中加热24 h以上;(5)待溶样弹冷却,开盖后置于140 ℃电热板上蒸干,然后加入1 mL HNO3并再次蒸干;(6)加入1 mL高纯HNO3、1 mL MQ水和1 mL内标In(浓度为1×10-6),再次将Teflon溶样弹放入钢套,拧紧后置于190 ℃烘箱中加热12 h以上;(7)将溶液转入聚乙烯瓶中,并用2% HNO3稀释至100 g以备ICP-MS测试。
-
本文涉及的地球化学数据见附件1,主要的地球化学指标见图2。Mo及MoEF变化趋势(图2a)相同,段Ⅰ分别在3.45×10-6~36.85×10-6、17.31~133.41变化。最大值(264.65×10-6、404.00)均在段Ⅱ出现,段Ⅲ稳定在较低值,分别在0.96×10-6~13.97×10-6、0.64~9.47变化。Th/U比值(图2b)平均值为1.13,段Ⅲ最高(3.32),段Ⅰ、段Ⅱ均较低(0.08~1.26)。Ce/Ce*(图2c)在段Ⅱ和段Ⅲ的变化稳定在0.77~0.99范围内,段Ⅰ平均值为0.64,并出现最小值(0.46)。Eu/Eu*(图2d)在段Ⅰ顶部突然增加,最大值达到1.78,段Ⅱ、段Ⅲ的变化范围分别为0.86~1.61、1.04~1.45。TOC含量(图2e)随岩性变化而变化。段Ⅰ部分较为波动,在0.20%~5.92%变化,平均值为2.29%。段Ⅱ的TOC含量最高(平均值为6.92%,最高值为12.48%)。段Ⅲ最低(0.14%~1.75%),平均值降至0.54%。TN含量的变化趋势(图2f)与TOC变化趋势几乎一致。段Ⅰ变化范围为0.03%~0.11%,段Ⅱ急剧上升,最大值为0.27%,平均值达到0.23%,段Ⅲ在0.07%~0.25%变化。δ13Corg值(图2g)从段Ⅰ至段Ⅲ逐渐增加,段Ⅰ变化范围为-31.51‰~-34.75‰,段Ⅱ在-30.99‰左右变化,段Ⅲ出现最高值(-27.43‰)。δ15N值(图2h)在段Ⅲ最低(-2.14‰),平均值为-1.27‰,而段Ⅰ最高(1.66‰),变化范围为-1.42‰~0.16‰,段Ⅱ相对稳定,在-0.43‰左右。

图 2 ZK0202井牛蹄塘组—变马冲组地球化学参数
Figure 2. The geochemical data from of well ZK0202 in the Niutitang Formation and Bianmachong Formation
其中,上述结果所涉及的计算方法如下:Mo元素富集因子(MoEF)计算公式为:MoEF=[(Mo/Al)sample]/[(Mo/Al)UCC],(Mo/Al)sample表示实测样品中的Mo与Al元素含量比,(Mo/Al)UCC表示上地壳中的Mo与Al元素含量比[35]。根据Shields et al.[36],Ce异常(Ce/Ce*)和Eu异常(Eu/Eu*)的计算公式分别为:Ce/Ce*=3CeN/(2LaN+NdN),Eu/Eu*=EuN/(SmN/GdN)1/2,其中XN表示利用后太古代平均页岩(PAAS)对元素浓度进行标准化[37]。
-
Mo是氧化还原敏感性元素之一,被广泛用于反映古海洋氧化还原条件[38]。Mo常以+6价的高价态形式溶解于氧化水体之中,在还原环境下将被还原为+4价的低价态,并保存在沉积物当中[39⁃40]。当水柱中出现游离态硫化氢时,Mo可在沉积物中显著富集[41⁃43],因此Mo含量及MoEF能够反映沉积期氧化还原环境。通常,Mo浓度超过100×10-6时代表持续的硫化环境,当浓度在(25~100)×10-6时代表间歇性硫化或低Mo浓度环境,当浓度小于25×10-6时代表非硫化环境[44]。如图2a所示,Mo在段Ⅰ内显示了中等的浓度和富集因子(平均值分别为21.26×10-6和79.89),说明研究区底水处于缺氧且间歇性硫化状态。在段Ⅱ内,Mo浓度和富集因子显著提升(平均值分别为100.3×10-6和100.2),指示研究区底水处于强硫化状态。段Ⅲ内Mo浓度和富集因子均值分别下降至6.81×10-6和4.45,暗示研究区底水处于次氧化—氧化状态。
Th/U比也是氧化还原重建的常用指标之一[45]。Th主要来自陆源,其化学性质较为稳定,受氧化还原条件和成岩作用影响较小。U作为变价元素,其化学性质受氧化还原影响较大,在缺氧条件下将出现明显富集。因此,Th/U比被认为能够较好地反映沉积期底水的氧化还原条件。一般地,Th/U比值小于2代表缺氧状态,2~7代表次氧化状态,大于8代表氧化状态[46]。如图2b所示,Th/U在段Ⅰ内均小于1,说明了缺氧的底水条件。Th/U在段Ⅱ内显示了最低的值(平均为0.32),说明缺氧状态在段Ⅱ内进一步加剧。段Ⅲ的Th/U显示了最高的平均值(约2.63),说明了较为氧化的底水条件。
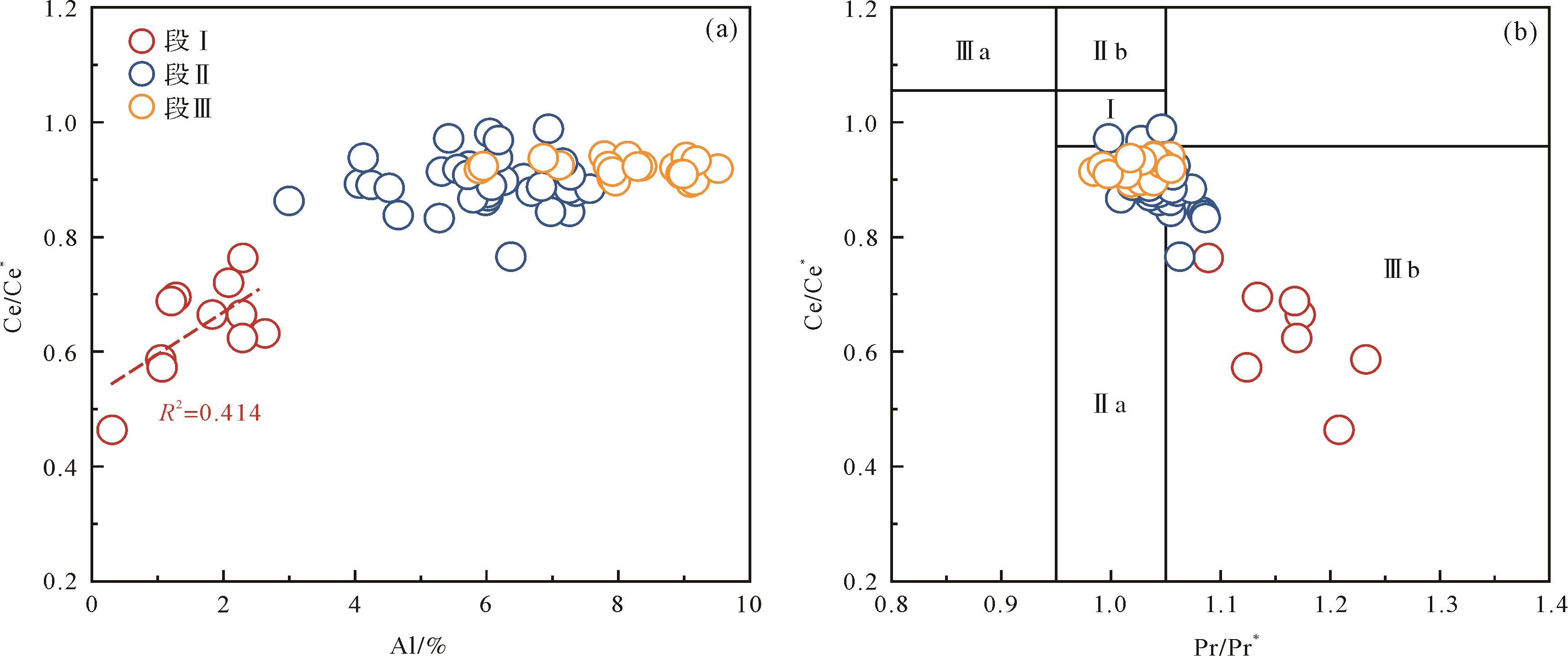
Ce有+3和+4两种价态,通常以+3价为主,不出现Ce的异常。当水体发生氧化时,Ce3+会被氧化为Ce4+,并与Mn4+发生类质同象替换而在铁锰氧化物中富集,从而造成海水呈现显著的负Ce异常。值得注意的是,由于陆源碎屑输入可能对样品的Ce异常产生重要影响,该指标更适合用于海水自生沉积岩(如碳酸盐岩、硅质岩、磷块岩等)。此外,利用Ce/Ce*-Pr/Pr*相关性图解可进一步判识由Pr异常造成的Ce异常假象[36]。如图3所示,本文硅质岩部分的Ce/Ce*和Al具有较好的相关性,碎屑岩部分(黑色页岩和粉砂岩)和Al显示了较弱的相关性(图3a)。这种现象说明硅质岩同时记录了来自海水和陆源的信息,而碎屑岩的Ce异常则主要受到陆源影响。因此,硅质岩更好地记录了海水的Ce异常信息。同时,硅质岩的Ce/Ce*全部位于Ce/Ce*-Pr/Pr*图解的Ⅲb区(图3b),说明硅质岩记录了真实的Ce异常[36]。因此,仅硅质岩部分的Ce异常可用于古氧化还原重建。如图2c所示,段Ⅰ的显示了显著的Ce负异常(平均值为0.64),暗示了硅质岩沉积期古海水处于次氧化状态。

图 3 PAAS标准化的Ce异常与(a)Al、(b)Pr异常相关性图
Figure 3. Correlation plot of Post⁃Archean Australian shale (PAAS)⁃normalized Ce anomalies with (a) Al and (b) Pr anomalies
Mo和Th/U指标总体上揭示了研究区一致的古氧化还原演化历程,即段Ⅰ缺氧(间歇性硫化)的底水环境向段Ⅱ持续硫化的底水环境演化,并进一步演化为次氧化—氧化的底水环境。上述古氧化还原演化历程和前人研究结果十分类似,均揭示了早寒武世(ca. 518 Ma)强硫化环境向氧化环境转换过程[8⁃9,47],反映了相对海平面变化对早寒武世华南古海洋氧化还原条件的整体控制[32]。与前人研究相比[8⁃9,32,47],Ce/Ce*指标的使用进一步揭示出华南早寒武世古海洋化变层相对位置的持续变化过程。结合Ce/Ce*、Mo和Th/U指标,可以推知早寒武世最早期(>526 Ma)化变层处于较深位置,而后逐渐向浅水移动(ca. 526~518 Ma),最后(<518 Ma)伴随着海退,化变层再次向深部移动,古海洋整体显示出更为氧化的状态[24]。
-
陆源物质的输入、早期成岩作用等都有可能改变沉积物中的氮含量,从而影响氮同位素组成[48⁃49]。因此,在讨论海洋氮循环之前,需要确定沉积物中氮同位素信号是否能够真实地反映原始海洋信息。
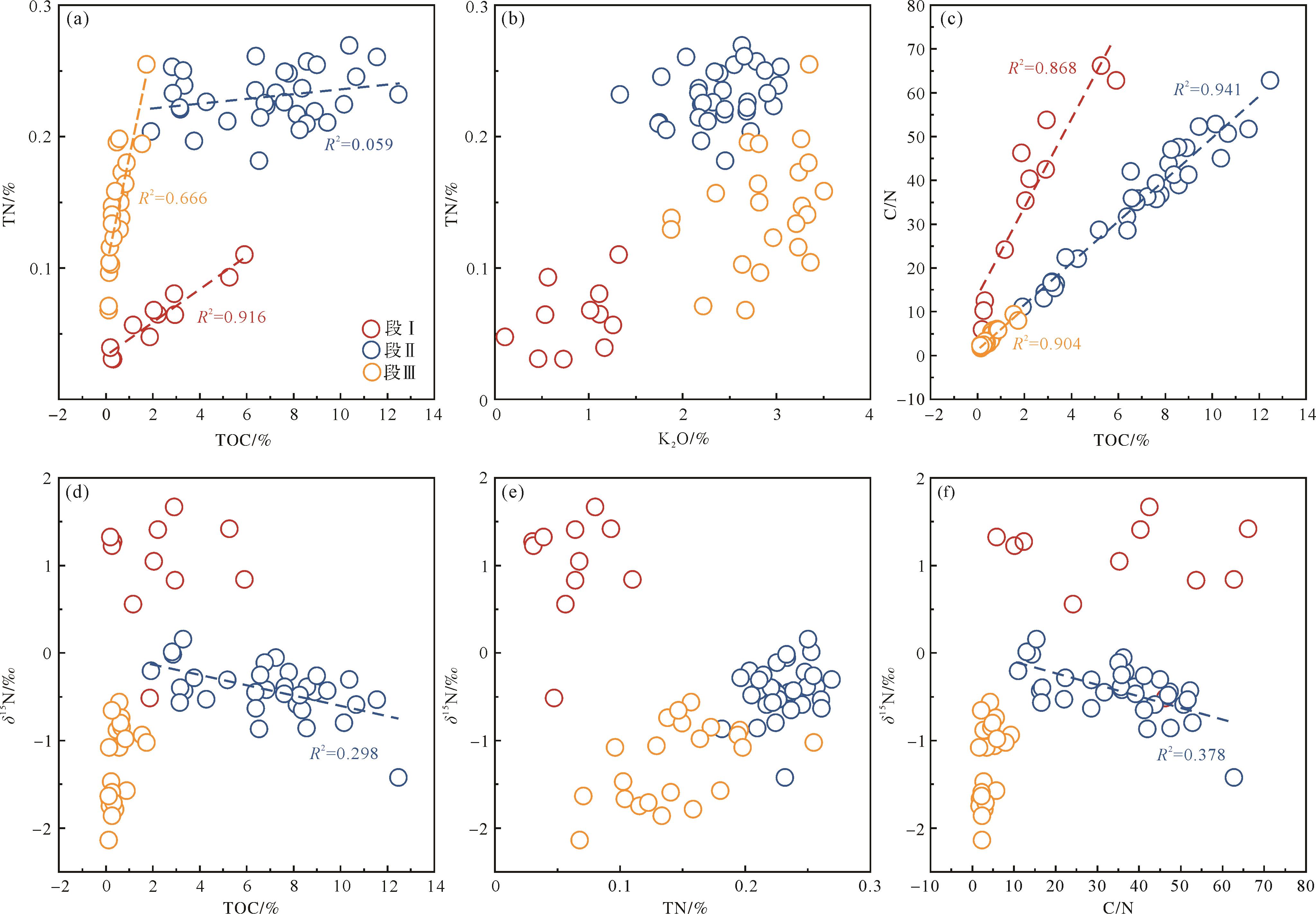
TOC和TN相关性是判断沉积物中氮来源的有效手段[16]。如图4a所示,段Ⅰ和段Ⅲ显示TOC和TN具有较好的线性相关性,说明硅质岩和粉砂质泥岩中氮主要以有机氮形态存在。因此,有机质是沉积物内最主要的氮来源[50]。在段Ⅱ内,TOC和TN未显示明显的线性相关性,似乎说明沉积物中氮并非以有机氮形态存在。然而该段内TN和K2O也没有显示线性相关性(图4b),说明陆源碎屑也并非沉积物中氮的主要来源。Chang et al.[9]将在水井沱组内观测到的上述现象解释为黏土结合氮丰度变化。因此,段Ⅱ内TOC和TN缺乏相关性也可能由该原因导致。此外,这种现象也可能说明了沉积物内氮同时赋存于有机质和黏土矿物中,因此破坏了有机质和总氮的相关性。这种猜测和TN轴上的非零截距是一致的(图4a)。

图 4 ZK0202井氮同位素信号有效性评估
Figure 4. Evaluation of the nitrogen isotope signal in well ZK0202
成岩作用是导致氮同位素变化的另一重要因素。通常,成岩作用中的氮损失可能造成C/N比的升高,进而导致TOC与C/N比显示出正相关性。因此,图4c显示研究样品皆受到了成岩作用影响。但是,δ15N-TOC、δ15N-TN、δ15N-C/N均没有表现出明显的线性相关性(图4d~f),这说明成岩作用并未对氮同位素产生显著影响。值得注意的是,段Ⅱ部分的δ15N-C/N、δ15N-TOC显示出了微弱的负相关关系。Chang et al.[9]认为该现象与有机质类型变化有关而非成岩作用所导致。
综上所述,尽管样品中的氮在沉积和成岩过程中可能产生了损失,但并没有显著影响样品的δ15N值。因此,ZK0202井样品的δ15N保留了原始沉积信息,可用于判断早寒武世研究区的氮循环过程。
-
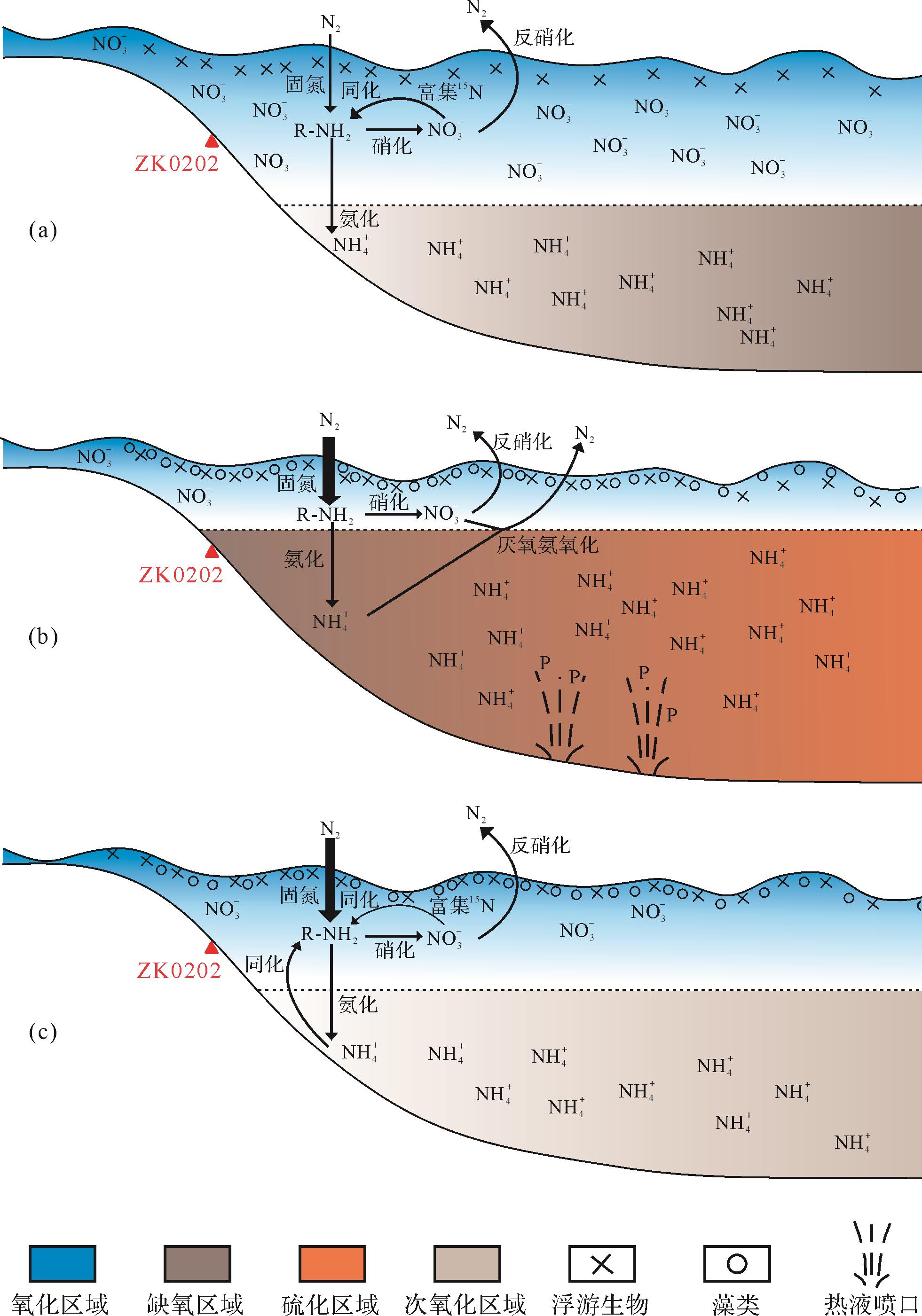
如图2h所示,段Ⅰ的δ15N的值皆为正值(平均值为1.01‰,最高值为1.66‰)。这样的氮同位素分布介于生物固氮作用(0‰)[51]和反硝化作用(5‰)[51]之间,说明此时的氮循环以生物固氮作用为主,同时有反硝化作用参与(图5a)。这种推测和氧化还原指标结果是完全一致的。由前文可知,段Ⅰ的古海洋氧化还原结构以较深化变层为特点的分层水体为主,这种氧化还原结构一方面有利于反硝化导致硝酸盐亏损,从而增强生物固氮作用;另一方面上部偏氧化水体也能使一部分残余硝酸盐被生物利用。从更长的时间尺度看,晚埃迪卡拉纪和早寒武幸运期均有较高的氮同位素值(分别为ca. 8.73‰[15]和ca. 8.8‰[52]),暗示了这些时期氮循环以反硝化作用为主。因此,硅质岩记录的氮同位素值说明了华南早寒武世古海洋在526 Ma之前就已经开始出现缺氧水体扩张迹象。

图 5 早寒武世生物与环境协同演化示意图
Figure 5. Schematic diagram of the co⁃evolution of early Cambrian organisms and the environment
-
在段Ⅱ内,δ15N整体体现为较为稳定的负值(平均值为-0.43‰)。这一结果与华南已报道同期氮同位素基本一致(-2‰~0‰)[47],说明固氮作用是同期华南扬子区氮循环的主要特征。前已述及,研究区在该段出现了明显的硫化扩大。而同期台地上金属硫化物矿层的广泛发育[21,27,53],富有机质烃源岩的广泛沉积[28,54]进一步揭示了硫化水体扩张并非局限于研究区,而是涉及整个上扬子台地。在这种条件下,强烈的反硝化将会导致大量的溶解性氮损失,进而形成以固氮作用为主导的氮循环模式(图5b)。
上述环境变化可能与同期热液活动密切相关,Xing et al.[55]认为这一时期广泛的磷块岩、金属硫化物沉积都是热液活动的结果。Eu异常同样证实了研究区受到了热液作用影响[56]。这种强烈的热液活动将释放大量的营养物,造成初级生产力提升和底部缺氧水体扩大。这种生产力水平的提升与段Ⅱ内有机碳同位素正偏是一致的(图2g)。虽然早寒武世逐渐正偏的碳同位素被归结为水柱氧化的结果[57⁃58],但这与氧化还原指标所展示的扩大缺氧水体并不相符(图2a,b)。因此,碳同位素的变化更可能说明了藻类有机质供给的大幅增加[59⁃61]。在这种条件下,强烈的氮需求也将进一步提升固氮作用,导致氮同位素值稳定在固氮作用范围内。由此可见,段Ⅱ内地球化学指标的系统变化揭示了地球深部对表生环境的重要影响。
-
段Ⅲ的氮同位素显示了进一步下降的特征(平均值为-1.27‰),暗示了氮循环依然以固氮作用为主。这一解释似乎和整体氧化的古海洋背景不符(图2a,b,e,g)。Wu et al.[32]提出这一现象与上涌引起的铵同化有关。但是相比较于Wu et al.[32]的结果,研究样品的氮同位素最低值(-2.14‰)明显偏高,并不完全支持铵同化作用。段Ⅲ内较低的氮同位素值可解释为段Ⅱ强烈反硝化作用的结果。由于前期溶解性氮库大量损失,加之生产者对氮源的大量需求,海洋中溶解性氮在段Ⅲ内依然没有被完全建立(图5c)。此外,次氧化水体中可能发生沉积物反硝化作用,造成水柱中短暂积累的硝酸盐损失,同样有利于固氮作用发生[62]。因此,虽然铵同化作用在研究区内可能发生,但固氮作用应该是该时期内最重要的氮循环途径。
-
通常认为,寒武纪生物爆发可分为前奏—序幕—主幕三个阶段[63]。其中,前奏主要以晚埃迪卡拉纪双胚层/类双胚层生物(即埃迪卡拉生物群)的出现为代表,序幕主要以早寒武世(ca. 539~518 Ma)小壳/遗迹化石的出现为代表,主幕则以ca. 518~505 Ma出现的高多样化后生动物(如澄江生物群和清江生物群)为代表[8⁃9,64⁃67]。本文主要涉及序幕和主幕两个阶段,且划分层段大致分别对应小壳化石出现(段Ⅰ)、灭绝(段Ⅱ)和澄江生物群出现(段Ⅲ)。研究结果显示,研究区段Ⅰ向段Ⅱ过渡时期发生了反硝化作用减少,固氮作用增强的现象,说明伴随着缺氧程度扩大,硝酸盐逐渐消耗殆尽。这种氧化还原变化和氮循环途径的转变在整个上扬子区都被广泛观察到[8,32,47,68],进一步暗示了这种转变是上扬子区的普遍特征。Wang et al.[8]基于这种变化提出硝酸盐浓度变化导致了小壳化石的出现和灭绝,这一解释是值得进一步商榷的。首先,上扬子区晚埃迪卡拉纪氮同位素值为整个埃迪卡拉—寒武纪转折期的最高值(ca. 8.73‰)[15],说明此时硝酸盐浓度最高,然而同期仅有埃迪卡拉生物群出现。其次,氮同位素值显示早寒武世幸运阶—第二阶中期硝酸盐浓度已逐渐下降,表明此时小壳化石的出现可能和硝酸盐并无直接联系。研究区段Ⅲ显示了氧化—次氧化环境,然而逐渐降低的氮同位素值说明此时硝酸盐浓度可能依然较低。这一时期华南最高的氮同位素值(ca. 6.1‰)仅出现在下扬子严家剖面[8],绝大多数上扬子区剖面氮同位素值依然显示出固氮作用的特征[47,68⁃69],指示澄江生物群出现时期可能并未出现普遍的硝酸盐浓度提升。这一推测进一步被清江生物群出现时期较低的氮同位素值(ca. -3‰)所证实[9]。这些现象或许均证实了硝酸盐浓度可能不是早寒武世生物演化的主要控制因素。由于硝酸盐浓度变化与氧化还原条件密切相关,其变化可能只是氧化还原的结果,氧化还原条件才是控制早寒武世生物演化的最重要因素。最后,由于目前对于硝酸盐浓度的解读主要基于氮同位素值,具有较强的多解性,未来模型的加入可能更好地量化不同阶段的硝酸盐浓度,最终确定其在早寒武世生物演化中的作用。
-
(1) 扬子东南缘斜坡区古海洋在第二阶—第三阶早期(ca. 526~518 Ma)发生了显著的硫化水体膨胀,而后逐渐向次氧化—氧化过渡(<518 Ma)。这些变化可能分别与热液引起的高初级生产力和海退有关。
(2) 斜坡区氮循环以固氮作用为主,在整个早寒武世并未发生明显变化,可能说明同期斜坡区硝酸盐浓度没有明显增加。
(3) 硝酸盐在生物大爆发中的作用有待进一步研究,而海洋氧化和食物链底层生物丰富可能是生物大爆发的重要诱因。
Early Cambrian Nitrogen Isotopic Characteristics and Its Significance for the Paleomarine Environment in the Southeastern Margin of Yangtze
-
摘要: 目的 早寒武世生物大爆发通常被认为与硝酸盐浓度增加有关,然而最新研究表明该时期硝酸盐浓度无明显变化。为查明硝酸盐对生物大爆发的影响,重建了这一时期古海洋环境与氮循环特征。 方法 以扬子东南缘斜坡区钻孔岩心(ZK0202井)为研究对象,通过对主量元素、微量元素、稀土元素、碳—氮同位素等地球化学指标进行实验分析。 结果 早寒武幸运阶—第二阶中期(>526 Ma)古海洋以深化变层的分层海洋为特征,同期固氮和反硝化作用都有发育,以固氮作用为主。第二阶晚期—第三阶早期(ca. 526~518 Ma)古海洋以强烈缺氧硫化为特征,固氮作用是最重要的氮循环途径。第三阶中晚期(<518 Ma)古海洋以次氧化—氧化为主,固氮作用依然发育。氧化还原状态的演化可能与海平面变化及热液作用引起的高初级生产力有关,而持续的固氮作用显示了同期硝酸盐浓度维持在较低水平。 结论 硝酸盐的多寡或许不是生物大爆发的主控因素,海洋氧化和丰富的食物链底层生物可能是生物大爆发的重要诱因。研究进一步提升了华南扬子东南缘早寒武世斜坡区古海洋环境与氮循环的研究程度,为正确认识该时期生物—环境协同演化机制提供了新的参考。Abstract: Objective The early Cambrian explosion is thought to be related to the increase of nitrate concentration; however, a recent study suggested that no significant change occurred in the nitrate concentration during this period. To identify the influence of nitrate on the biological explosion, this study reconstructed the characteristics of the paleomarine environment and nitrogen cycle during this period. Methods The borehole core (well ZK0202) in the slope area of the southeast margin of the Yangtze was used as the anatomical research object, and the major elements, trace elements, rare earth elements, carbon-nitrogen isotopes, and other indices were analyzed. Results The early Cambrian Fortunian-middle Age 2 (>526 Ma) paleomarine environment was characterized by a stratified ocean with a deepened chemocline, and both nitrogen fixation and denitrification developed during the same period, with nitrogen fixation being the dominant effect. In the late Age 2-early Age 3 (ca. 526-518 Ma), the paleomarine environment was characterized by strong anoxic-euxinic behavior, and nitrogen fixation was the most important nitrogen cycle pathway. The middle and late Age 3 (<518 Ma) paleomarine environment was dominated by suboxic-oxic behavior, and nitrogen fixation was still developed. The evolution of the redox state may be related to the high primary productivity induced by hydrothermal action, and the persistent nitrogen fixation shows that the nitrate concentration was maintained at a low level during the same period. Conclusions Therefore, the abundance of nitrate may not be the main controlling factor for the biological explosion, and ocean oxidation and the abundance of organisms at the base of the food chain may be important triggers. The above research results further enhance the degree of research on the paleomarine environment and nitrogen cycle in the early Cambrian slope area of the southeast margin of the South China Yangtze, providing new references for the correct understanding of the environment-biological co-evolution in this period.
-
Key words:
- early Cambrian /
- redox /
- nitrogen cycle /
- biological explosion
注释:1) 脚注:1)http://www.cjxb.ac.cn/cn/article/doi/10.14027/j.issn.1000-0550.2023.130. -
图 1 (a)寒武纪早期全球古地理图(据文献[19⁃20]修改);(b)寒武纪早期扬子台地古地理图(据文献[23]修改);(c)ZK0202井牛蹄塘组—变马冲组岩性柱
Figure 1. (a) Early Cambrian global paleogeography (modified from references [19⁃20]); (b) paleogeographic map of the Yangtze Platform during the early Cambrian (modified from reference [23]; (c) lithological column of the Niutitang Formationand Bianmachong Formation in well ZK0202)
图 2 ZK0202井牛蹄塘组—变马冲组地球化学参数
red, blule, and yellow cycles represent sections Ⅰ, Ⅱ, and Ⅲ, respectively
Figure 2. The geochemical data from of well ZK0202 in the Niutitang Formation and Bianmachong Formation
Fig.2
图 3 PAAS标准化的Ce异常与(a)Al、(b)Pr异常相关性图
field Ⅰ, no anomaly; field Ⅱa, positive La anomaly causes apparent negative Ce anomaly; field Ⅱb, negative La anomaly causes apparent positive Ce anomaly;field Ⅲa, real positive Ce anomaly; and field Ⅲb, real negative Ce anomaly
Figure 3. Correlation plot of Post⁃Archean Australian shale (PAAS)⁃normalized Ce anomalies with (a) Al and (b) Pr anomalies
Fig.3
图 4 ZK0202井氮同位素信号有效性评估
(a) TOC vs. TN; (b) K2O vs. TN; (c) TOC vs. C/N; (d) TOC vs. δ15N; (e) TN vs. δ15N; (f) C/N vs. δ15N
Figure 4. Evaluation of the nitrogen isotope signal in well ZK0202
Fig.4 -
[1] Knoll A H, Carroll S B. Early animal evolution: Emerging views from comparative biology and geology[J]. Science, 1999, 284(5423): 2129-2137. [2] Amthor J E, Grotzinger J P, Schröder S, et al. Extinction of cloudina and namacalathus at the Precambrian-Cambrian boundary in Oman[J]. Geology, 2003, 31(5): 431-434. [3] Scott C, Lyons T W, Bekker A, et al. Tracing the stepwise oxygenation of the Proterozoic ocean[J]. Nature, 2008, 452(7186): 456-459. [4] Chen X, Ling H F, Vance D, et al. Rise to modern levels of ocean oxygenation coincided with the Cambrian radiation of animals[J]. Nature Communications, 2015, 6: 7142. [5] Sahoo S K, Planavsky N J, Jiang G, et al. Oceanic oxygenation events in the anoxic Ediacaran ocean[J]. Geobiology, 2016, 14(5): 457-468. [6] Och L M, Shields-Zhou G A, Poulton S W, et al. Redox changes in early Cambrian black shales at Xiaotan section, Yunnan pro-vince, South China[J]. Precambrian Research, 2013, 225: 166-189. [7] Pi D H, Liu C Q, Shields-Zhou G A, et al. Trace and rare earth element geochemistry of black shale and kerogen in the early Cambrian Niutitang Formation in Guizhou province, South China: Constraints for redox environments and origin of metal enrichments[J]. Precambrian Research, 2013, 225: 218-229. [8] Wang D, Ling H F, Struck U, et al. Coupling of ocean redox and animal evolution during the Ediacaran-Cambrian transition[J]. Nature Communications, 2018, 9(1): 2575. [9] Chang C, Wang Z F, Huang K J, et al. Nitrogen cycling during the peak Cambrian explosion[J]. Geochimica et Cosmochimica Acta, 2022, 336: 50-61. [10] Ader M, Thomazo C, Sansjofre P, et al. Interpretation of the nitrogen isotopic composition of Precambrian sedimentary rocks: Assumptions and perspectives[J]. Chemical Geology, 2016, 429: 93-110. [11] Chen Y, Diamond C W, Stüeken E E, et al. Coupled evolution of nitrogen cycling and redoxcline dynamics on the Yangtze Block across the Ediacaran-Cambrian transition[J]. Geochimica et Cosmochimica Acta, 2019, 257: 243-265. [12] Anbar A D, Knoll A H. Proterozoic ocean chemistry and evolution: A bioinorganic bridge?[J]. Science, 2002, 297(5584): 1137-1142. [13] Anbar A D, Duan Y, Lyons T W, et al. A whiff of oxygen before the Great Oxidation Event?[J]. Science, 2007, 317(5846): 1903-1906. [14] Ader M, Sansjofre P, Halverson G P, et al. Ocean redox structure across the Late Neoproterozoic oxygenation event: A nitrogen isotope perspective[J]. Earth and Planetary Science Letters, 2014, 396: 1-13. [15] Cremonese L, Shields-Zhou G A, Struck U, et al. Nitrogen and organic carbon isotope stratigraphy of the Yangtze Platform during the Ediacaran-Cambrian transition in South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 398: 165-186. [16] Kikumoto R, Tahata M, Nishizawa M, et al. Nitrogen isotope chemostratigraphy of the Ediacaran and early Cambrian platform sequence at Three Gorges, South China[J]. Gondwana Research, 2014, 25(3): 1057-1069. [17] Li C, Cheng M, Zhu M Y, et al. Heterogeneous and dynamic marine shelf oxygenation and coupled early animal evolution[J]. Emerging Topics in Life Sciences, 2018, 2(2): 279-288. [18] 杨桦. 埃迪卡拉纪—寒武纪之交南华盆地深水氧化还原状态[D]. 贵阳:贵州师范大学,2023:13-122. Yang Hua. Redox condition of deepwater in the Nanhua Basin at the Ediacaran-Cambrian transition[D]. Guiyang: Guizhou Normal University, 2023: 13-122. [19] Li C, Zhang Z H, Jin C S, et al. Spatiotemporal evolution and causes of marine euxinia in the early Cambrian Nanhua Basin (South China)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 546: 109676. [20] Zhang S H, Li H Y, Jiang G Q, et al. New paleomagnetic results from the Ediacaran Doushantuo Formation in South China and their paleogeographic implications[J]. Precambrian Research, 2015, 259: 130-142. [21] Wang J, Li Z X. History of Neoproterozoic rift basins in South China: Implications for Rodinia break-up[J]. Precambrian Research, 2003, 122(1/2/3/4): 141-158. [22] 王剑,刘宝珺,潘桂棠. 华南新元古代裂谷盆地演化:Rodinia超大陆解体的前奏[J]. 矿物岩石,2001,21(3):135-145. Wang Jian, Liu Baojun, Pan Guitang. Neoproterozoic rifting history of South China significance to Rodinia breakup[J]. Journal of Mineralogy and Petrology, 2001, 21(3): 135-145. [23] Goldberg T, Strauss H, Guo Q J, et al. Reconstructing marine redox conditions for the early Cambrian Yangtze Platform: Evidence from biogenic sulphur and organic carbon isotopes[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2007, 254(1/2): 175-193. [24] Jin C S, Li C, Algeo T J, et al. A highly redox-heterogeneous ocean in South China during the early Cambrian (~529-514 Ma): Implications for biota-environment co-evolution[J]. Earth and Planetary Science Letters, 2016, 441: 38-51. [25] Jiang G Q, Wang X Q, Shi X Y, et al. The origin of decoupled carbonate and organic carbon isotope signatures in the early Cambrian (ca. 542–520 Ma) Yangtze Platform[J]. Earth and Planetary Science Letters, 2012, 317-318: 96-110. [26] 陈代钊,汪建国,严德天,等. 中扬子地区早寒武世构造—沉积样式与古地理格局[J]. 地质科学,2012,47(4):1052-1070. Chen Daizhao, Wang Jianguo, Yan Detian, et al. Tectono-depositional patterns and palaeogeography in the Middle Yangtze River region during the early Cambrian[J]. Chinese Journal of Geology, 2012, 47(4): 1052-1070. [27] Wang J G, Chen D Z, Yan D T, et al. Evolution from an anoxic to oxic deep ocean during the Ediacaran-Cambrian transition and implications for bioradiation[J]. Chemical Geology, 2012, 306-307: 129-138. [28] Chen D Z, Zhou X Q, Fu Y, et al. New U-Pb zircon ages of the Ediacaran-Cambrian boundary strata in South China[J]. Terra Nova, 2015, 27(1): 62-68. [29] 金承胜,李超,彭兴芳,等. 华南寒武纪早期海洋化学状态的时空波动[J]. 中国科学:地球科学,2014,44(5):851-863. Jin Chengsheng, Li Chao, Peng Xingfang, et al. 2014. Spatiotemporal variability of ocean chemistry in the early Cambrian, South China[J]. Science China: Earth Sciences, 2014, 44(5): 851-863. [30] Wang D, Struck U, Ling H F, et al. Marine redox variations and nitrogen cycle of the early Cambrian southern margin of the Yangtze Platform, South China: Evidence from nitrogen and organic carbon isotopes[J]. Precambrian Research, 2015, 267: 209-226. [31] Xu L G, Lehmann B, Mao J W, et al. Re-Os age of polymetallic Ni–Mo–PGE-Au mineralization in early Cambrian black shales of South China: A reassessment[J]. Economic Geology, 2011, 106(3): 511-522. [32] Wu Y W, Tian H, Jia W L, et al. Nitrogen isotope evidence for stratified ocean redox structure during Late Ediacaran to Cambrian Age 3 in the Yangtze Block of South China[J]. Chemical Geology, 2022, 589: 120679. [33] 朱茂炎,孙智新,杨爱华,等. 中国寒武纪岩石地层划分和对比[J]. 地层学杂志,2021,45(3):223-249. Zhu Maoyan, Sun Zhixin, Yang Aihua, et al. Lithostratigraphic subdivision and correlation of the Cambrian in China[J]. Journal of Stratigraphy, 2021, 45(3): 223-249. [34] Yang C, Li X H, Zhu M Y, et al. Geochronological constraint on the Cambrian Chengjiang biota, South China[J]. Journal of the Geological Society, 2018, 175(4): 659-666. [35] McLennan S M. Relationships between the trace element composition of sedimentary rocks and upper continental crust[J]. Geochemistry, Geophysics, Geosystems, 2001, 2(4): 2000GC0 00109. [36] Shields G, Stille P. Diagenetic constraints on the use of cerium anomalies as palaeoseawater redox proxies: An isotopic and REE study of Cambrian phosphorites[J]. Chemical Geology, 2001, 175(1/2): 29-48. [37] McLennan S M. Rare earth elements in sedimentary rocks; influence of provenance and sedimentary processes[J]. Reviews in Mineralogy and Geochemistry, 1989, 21(1): 169-200. [38] Tribovillard N, Algeo T J, Lyons T, et al. Trace metals as paleoredox and paleoproductivity proxies: An update[J]. Chemical Geology, 2006, 232(1/2): 12-32. [39] 张明亮,郭伟,沈俊,等. 古海洋氧化还原地球化学指标研究新进展[J]. 地质科技情报,2017,36(4):95-106. Zhang Mingliang, Guo Wei, Shen Jun, et al. New progress on geochemical indicators of ancient oceanic redox condition[J]. Geological Science and Technology Information, 2017, 36(4): 95-106. [40] Algeo T J, Li C. Redox classification and calibration of redox thresholds in sedimentary systems[J]. Geochimica et Cosmochimica Acta, 2020, 287: 8-26. [41] Helz G R, Miller C V, Charnock J M, et al. Mechanism of molybdenum removal from the sea and its concentration in black shales: EXAFS evidence[J]. Geochimica et Cosmochimica Acta, 1996, 60(19): 3631-3642. [42] Piper D Z, Perkins R B. A modern vs. Permian black shale: The hydrography, primary productivity, and water-column chemistry of deposition[J]. Chemical Geology, 2004, 206(3/4): 177-197. [43] Piper D Z, Calvert S E. A marine biogeochemical perspective on black shale deposition[J]. Earth-Science Reviews, 2009, 95(1/2): 63-96. [44] Scott C, Lyons T W. Contrasting molybdenum cycling and isotopic properties in euxinic versus non-euxinic sediments and sedimentary rocks: Refining the paleoproxies[J]. Chemical Geology, 2012, 324-325: 19-27. [45] 常华进,储雪蕾,黄晶,等. 埃迪卡拉纪末期华南盆地深水环境的微量元素约束[J]. 兰州大学学报(自然科学版),2011,47(5):17-23,29. Chang Huajin, Chu Xuelei, Huang Jing, et al. Trace element constraints on deep-water redox conditions of the terminal Ediacaran in South China Basin[J]. Journal of Lanzhou University (Natural Sciences), 2011, 47(5): 17-23,29. [46] Wignall P B, Twitchett R J. Oceanic anoxia and the end Permian mass extinction[J]. Science, 1996, 272(5265): 1155-1158. [47] Liu Y, Magnall J M, Gleeson S A, et al. Spatio-temporal evolution of ocean redox and nitrogen cycling in the early Cambrian Yangtze ocean[J]. Chemical Geology, 2020, 554: 119803. [48] Robinson R S, Kienast M, Albuquerque A L, et al. A review of nitrogen isotopic alteration in marine sediments[J]. Paleoceanography, 2012, 27(4): PA4203. [49] Stüeken E E, Kipp M A, Koehler M C, et al. The evolution of Earth's biogeochemical nitrogen cycle[J]. Earth-Science Reviews, 2016, 160: 220-239. [50] Calvert S E. Beware intercepts: Interpreting compositional ratios in multi-component sediments and sedimentary rocks[J]. Organic Geochemistry, 2004, 35(8): 981-987. [51] Sigman D M, Karsh K L, Casciotti K L. Nitrogen isotopes in the ocean[M]//Steele J H, Thorpe S A, Turekian K K. Encyclopedia of ocean sciences. 2nd ed. San Diego: Academic Press, 2009: 40-54. [52] Cremonese L, Shields-Zhou G, Struck U, et al. Marine biogeochemical cycling during the early Cambrian constrained by a nitrogen and organic carbon isotope study of the Xiaotan section, South China[J]. Precambrian Research, 2013, 225: 148-165. [53] Guo Q J, Shields G A, Liu C Q, et al. Trace element chemostratigraphy of two Ediacaran–Cambrian successions in South China: Implications for organosedimentary metal enrichment and silicification in the early Cambrian[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2007, 254(1/2): 194-216. [54] Xu L G, Lehmann B, Mao J W, et al. Mo isotope and trace element patterns of lower Cambrian black shales in South China: Multi-proxy constraints on the paleoenvironment[J]. Chemical Geology, 2012, 318-319: 45-59. [55] Xing J Q, Jiang Y H, Xian H Y, et al. Hydrothermal activity during the formation of REY-rich phosphorites in the early Cambrian Gezhongwu Formation, Zhijin, South China: A micro- and nano-scale mineralogical study[J]. Ore Geology Reviews, 2021, 136: 104224. [56] 赵瞻,谢渊,刘建清,等. 渝东南下寒武统黑色岩系稀土元素地球化学特征与沉积环境[J]. 沉积与特提斯地质,2011,31(2):49-54. Zhao Zhan, Xie Yuan, Liu Jianqing, et al. REE geochemical signatures and sedimentary environments of the lower Cambrian black rock series in southeastern Chongqing and its adjacent areas[J]. Sedimentary Geology and Tethyan Geology, 2011, 31(2): 49-54. [57] Wang N, Li M J, Tian X W, et al. Climate-ocean control on the depositional watermass conditions and organic matter enrichment in lower Cambrian black shale in the Upper Yangtze Platform[J]. Marine and Petroleum Geology, 2020, 120: 104570. [58] 曹光耀,刘宇,侯明才,等. 川南威远地区早寒武世氮循环及古环境意义[J]. 沉积学报,2024,42(6):2031-2041. Cao Guangyao, Liu Yu, Hou Mingcai, et al. Nitrogen cycle and paleoenvironmental implications in the Weiyuan area, southern Sichuan during the early Cambrian[J]. Acta Sedimentologica Sinica, 2024, 42(6): 2031-2041. [59] 何洪茜,肖加飞,杨海英,等. 黔北地区下寒武统牛蹄塘组泥岩沉积环境及页岩气勘探潜力[J]. 沉积与特提斯地质,2024,44(2):267-277. He Hongxi, Xiao Jiafei, Yang Haiying, et al. Sedimentary environment and shale gas exploration potential of lower Cambrian Niutitang Formation in northern Guizhou[J]. Sedimentary Geology and Tethyan Geology, 2024, 44(2): 267-277. [60] 赵建华,金之钧,林畅松,等. 上扬子地区下寒武统筇竹寺组页岩沉积环境[J]. 石油与天然气地质,2019,40(4):701-715. Zhao Jianhua, Jin Zhijun, Lin Changsong, et al. Sedimentary environment of the lower Cambrian Qiongzhusi Formation shale in the Upper Yangtze region[J]. Oil & Gas Geology, 2019, 40(4): 701-715. [61] 祝庆敏,卢龙飞,潘安阳,等. 湘西地区下寒武统牛蹄塘组页岩沉积环境与有机质富集[J]. 石油实验地质,2021,43(5):797-809,854. Zhu Qingmin, Lu Longfei, Pan Anyang, et al. Sedimentary environment and organic matter enrichment of the lower Cambrian Niutitang Formation shale, western Hunan pro-vince, China[J]. Petroleum Geology & Experiment, 2021, 43(5): 797-809, 854. [62] 朱艳宸,李丽,王鹏,等. 海洋氮循环中稳定氮同位素变化与地质记录研究进展[J]. 地球科学进展,2020,35(2):167-179. Zhu Yanchen, Li Li, Wang Peng, et al. Progress in the study of marine stable nitrogen isotopic changes and its geological records[J]. Advances in Earth Science, 2020, 35(2): 167-179. [63] 舒德干. 寒武纪大爆发与动物树的成型[J]. 地球科学与环境学报,2009,31(2):111-134. Shu Degan. Cambrian explosion: Formation of tree of animals[J]. Journal of Earth Sciences and Environment, 2009, 31(2): 111-134. [64] Steiner M, Zhu M Y, Zhao Y L, et al. Lower Cambrian Burgess Shale-type fossil associations of South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 220(1/2): 129-152. [65] Steiner M, Li G X, Qian Y, et al. Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2007, 254(1/2): 67-99. [66] 朱茂炎. 动物的起源和寒武纪大爆发:来自中国的化石证据[J]. 古生物学报,2010,49(3):269-287. Zhu Maoyan. The origin and Cambrian explosion of animals: Fossil evidences from China[J]. Acta Palaeontologica Sinica, 2010, 49(3): 269-287. [67] Hammarlund E U, Gaines R R, Prokopenko M G, et al. Early Cambrian oxygen minimum zone-like conditions at Chengjiang[J]. Earth and Planetary Science Letters, 2017, 475: 160-168. [68] Wang H Z, Wang D, Wei G Y, et al. Increases in marine environmental heterogeneity during the early animal innovations: Evidence from nitrogen isotopes in South China[J]. Precambrian Research, 2022, 369: 106501. [69] Xu D T, Wang X Q, Shi X Y, et al. Nitrogen cycle perturbations linked to metazoan diversification during the early Cambrian[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2020, 538: 109392. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 190
- HTML全文浏览量: 30
- PDF下载量: 27
- 被引次数: 0

