
HTML
-
在过去三十年里,“carbonate factory(碳酸盐工厂/碳酸盐生产工厂)”的定义不断被完善,沉积环境、沉积物可容空间,以及生物作用等因素对碳酸盐生产的影响逐渐受到研究者重视。关于碳酸盐沉积物的生产过程,James et al.[1⁃2]用“Carbonates are born,not made”形象地表达了碳酸盐沉积物的生物成因属性,将其描述为温暖浅水环境中由于生物骨骼钙化或者海水化学沉淀而导致碳酸盐沉积物“出生”的过程,并强调它是具有较高的生物生产率的浅水沉积区。Tucker et al.[3]认为在水深15 m之内高碳酸盐产率的潮下带是碳酸盐沉积物生产的主要场所;Schlager[4]认为它不仅是碳酸盐沉积物的生产场所,还是识别、区分碳酸盐沉积过程和分类方案的载体;Pomar et al. [5]指出碳酸盐工厂除了是碳酸盐沉积物生产场所外,在识别工厂建造者、不同时期碳酸盐岩沉积内在特异性差别上具有独特价值。碳酸盐工厂的形成受到不同生物种类、沉淀方式、水体环境(水深、水动力、盐度、光照等)、营养水平、气候条件(温度等)、海洋化学及陆源输入等因素的影响,根据其形成条件可以被划分为不同的工厂类型[6⁃8]。目前运用最广泛的分类方案主要有两种:一类是Schlager[4,7]的方案,通过主要的底栖生物种类,将碳酸盐工厂分为“热带或热带浅水工厂”(tropical或tropical shallow-water)、“温凉水工厂”(cool water)和“灰泥丘工厂”(mud-mound)三种主要形式。另一类碳酸盐工厂体系是由Pomar et al.[5]提出的,强调各种物理和生态因素对碳酸盐工厂发育过程的调节,还强调了多种因素对其影响(如营养水平、水体能量、温度、透光度等),该方案主要强调了碳酸盐建造过程中的主要生产者,并且认为碳酸盐工厂的发展是一个从简单到复杂、从低级到高级的过程。Reijmer[9]对碳酸盐生产工厂概念的早期发展历史进行了全面回顾,并对Schlager[4,7]的碳酸盐生产工厂分类中不同工厂的特点进行补充,并在之前的分类基础上增加了冷水珊瑚工厂和浮游生物工厂,同时指出每个工厂均有独特的特征,例如生产窗口、沉积物生产和运输、沉积体形态和坡度,正是这些独特的变量标志着每个工厂对不同尺度下环境变化的综合响应。综上所述,准确识别不同碳酸盐工厂,阐明其控制因素,厘清它们的演化过程,可以作为了解碳酸盐岩沉积系统演化的绝佳切入点。
Reijmer[9]分类方案中的热带浅水工厂,主要指分布于南北纬30°内的热带和亚热带,温暖、光照充足、营养贫乏但含氧量高的海洋表层环境;虽然热带浅水碳酸盐工厂分布区域局限,但碳酸盐生产率最高[10]。一般认为热带浅水工厂的主要建造者为光合自养绿藻类以及直接共生的珊瑚、有孔虫和某些软体动物,通过光合作用、生物矿化作用以及有机矿化作用实现碳酸盐组分的形成和保存。地质历史时期的热带浅水碳酸盐生产工厂的建造者可能更为丰富,常见类型包括内碎屑、生物碎屑(骨屑)和非生物碎屑(非骨屑,如鲕粒、似球粒和核形石等)[11];一些环境因素,如水动力条件、营养水平、温度及碳酸盐矿物饱和度等因素,对不同类型碳酸盐颗粒的形成和保存起到了决定性影响[12⁃14]。而不同的碳酸盐颗粒组合可能代表了各自特异化的碳酸盐生产条件和生产模式,即细分的碳酸盐工厂类型。碳酸盐生产工厂可能包含了该沉积环境中共存或交替出现的多种特异化碳酸盐生产过程。前人在进行碳酸盐生产工厂的研究时多关注同一沉积环境中的整体生产模式,所以分析得出的工厂分类通常与相应的沉积相吻合,而对于同一沉积环境中更细分的、特异化工厂的研究则较为薄弱。本文旨在利用沉积学分析方法识别出热带浅水生产工厂中细分的特异化工厂类型,分析其形成和发育的环境,探索不同细分工厂转换的调控因素,特别是针对碳酸盐工厂沉积学特征的定量化表征方法进行进一步的尝试,为深入理解和描述不同类型碳酸盐工厂的组成特征、作用机制以及控制因素提供可借鉴的方案。
四川盆地西北部绵竹汉旺地区的马鞍塘组沉积序列较完整,碳酸盐岩沉积体系中详细地记录了该时期碳酸盐工厂转换和碳酸盐台地消亡的整个过程[15⁃19]。基于前人对川西北马鞍塘组沉积特征的演化,以及长时间尺度上极端事件影响碳酸盐工厂发育过程的研究[20⁃25],本文针对该区域马鞍塘组中不同颗粒组合所代表的特异化细分工厂进行进一步的分类,并进行精细化、定量化的沉积学及统计学分析,探讨短时间尺度内不同细分工厂的分类、特征及转变的驱动因素。
-
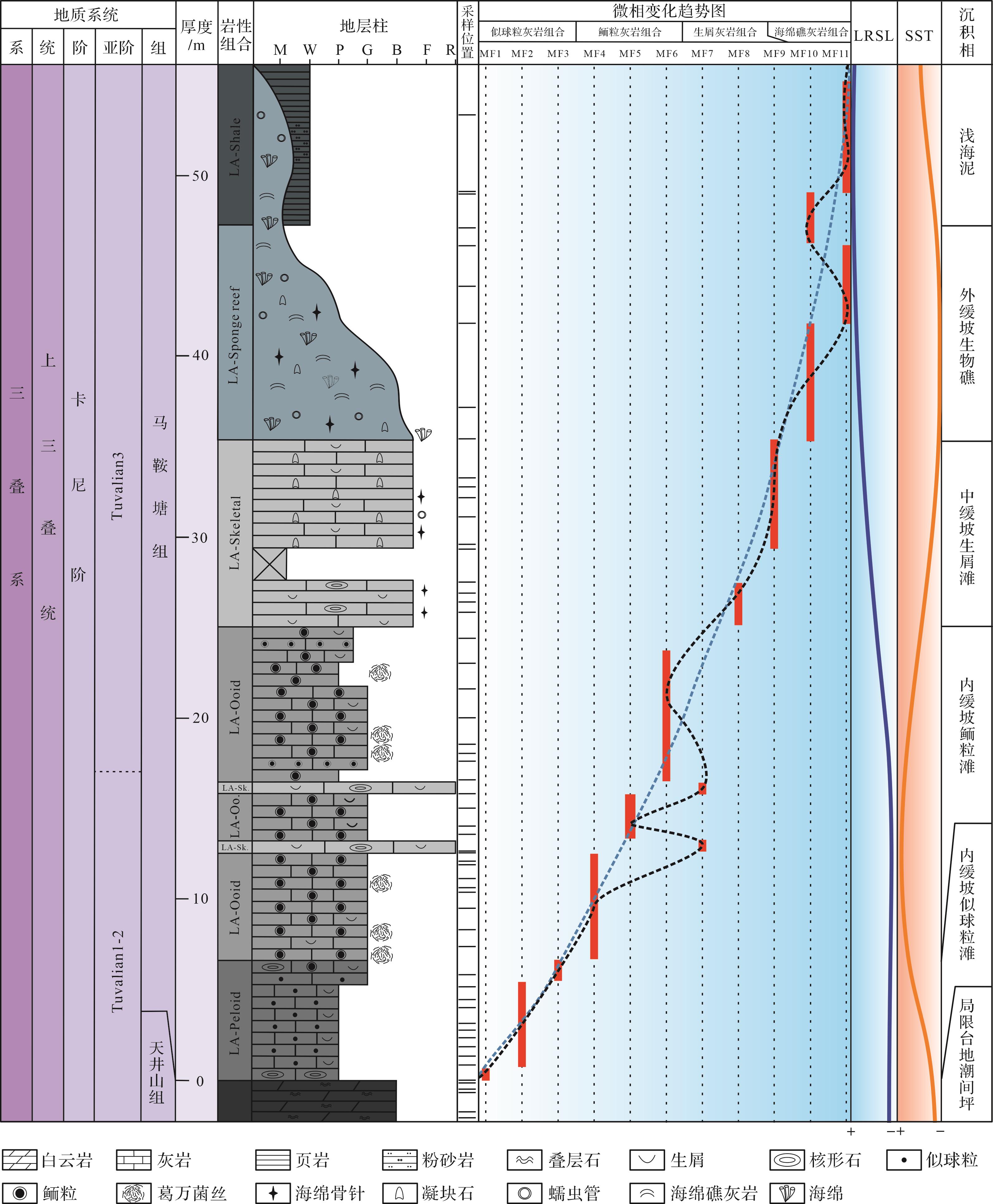
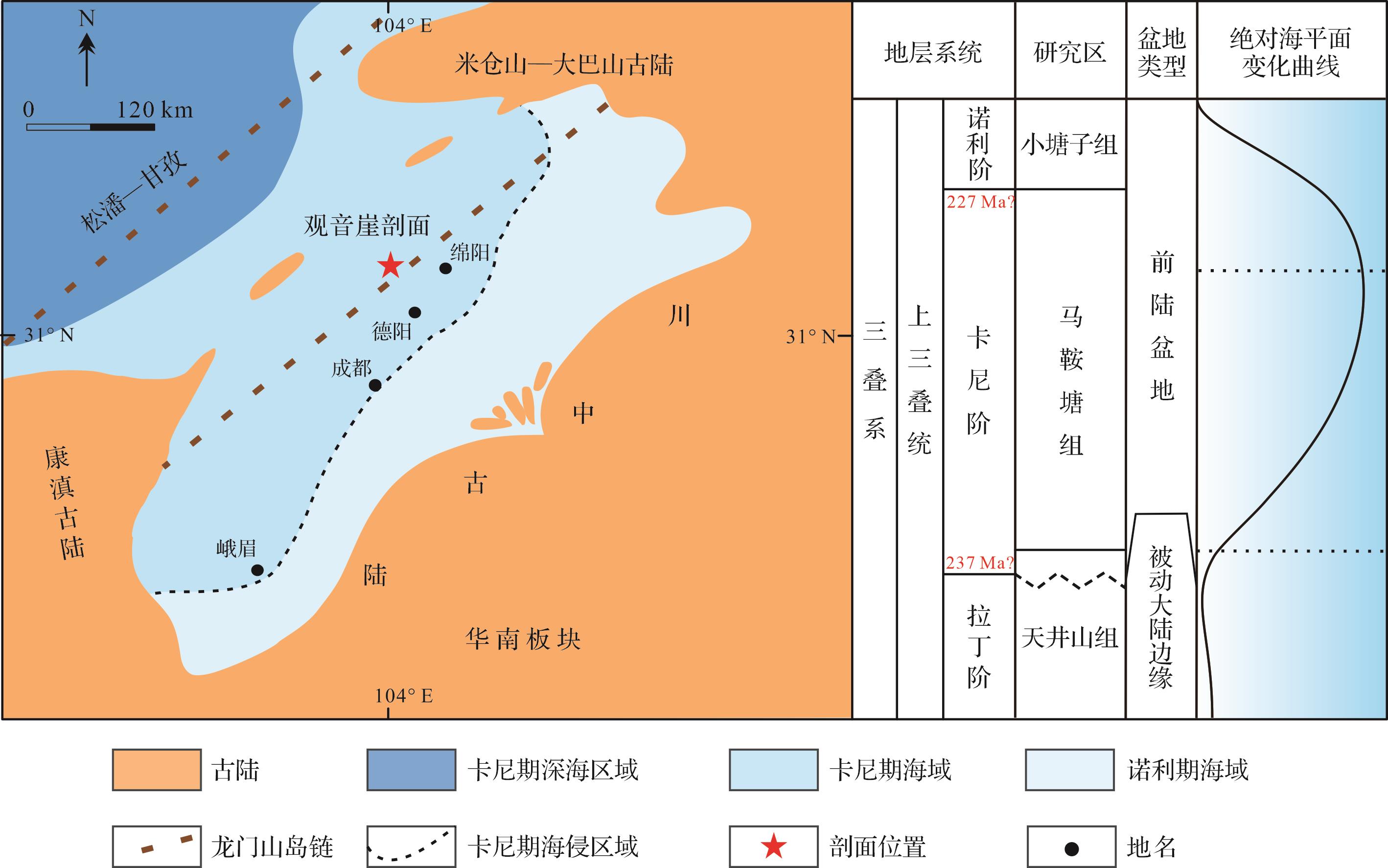
中—晚三叠世,全球板块运动频繁,位于东特提斯洋的四川盆地也受到了印支运动的影响,羌塘板块、华北板块和扬子板块发生汇聚和碰撞,导致了古特提斯洋支盆的金沙江缝合带闭合,形成了一系列北东向隆起与坳陷,如龙门山断续隆起、康滇隆起及西北坳陷区[26⁃27]。此时四川盆地西北部被龙门山岛链环绕,隔断了松潘—甘孜地块;北部受米仓山—大巴山古陆的限制;西南部被康滇古陆阻隔;东南部为川中古陆,受控于华南造山带隆起,有着“三隆围一坳”的格局(图1)[18,21,28⁃29]。晚三叠世,四川盆地西北部经历多次海侵与海退,进入海陆转换的重要时期[30⁃31]。在晚三叠世早期,松潘—甘孜地区仍为深海沉积,为一残留海盆,龙门山断裂带将甘孜深海盆地与川西盆地分隔开来[32⁃34]。随后古特提斯洋海水越过龙门山岛链,在研究区形成了一个半封闭的海湾,在广元—江油—绵竹一带广泛发育海相碳酸盐岩沉积[35⁃36]。同时在该时期卡尼期潮湿气候事件达到高潮,降雨量的增加导致大陆风化加重,大量的陆源营养物质或有机物输入,为富有机质页岩的产生提供了良好的物质基础[37⁃38]。研究区最后一套碳酸盐岩沉积完整地记录了晚三叠世卡尼期从海相沉积到陆相沉积的转变过程[15]。

Figure 1. Late Triassic Carnian paleogeography and lithostratigraphic column of the western Sichuan Basin (modified from references [18,21,28⁃29])
观音崖剖面位于四川省绵竹市汉旺镇北约2 km处(31°28′10.32" N, 104°08′49.52" E),剖面沿绵远河两岸展布。野外描述始于中三叠统拉丁阶天井山组的顶部,止于上三叠统卡尼期马鞍塘组的上部,整个剖面在绵远河的两岸均有出露,地层出露良好。马鞍塘组地层厚度为30~70 m,与其上覆的小塘子组和下伏的天井山组在研究区出露广泛[39⁃40]。碳酸盐岩沉积序列自下往上多期的岩性变化,马鞍塘组主要为似球粒灰岩、鲕粒灰岩、生屑灰岩、生物礁灰岩,以及覆盖在其上的薄—中层泥页岩。上覆的小塘子组为灰色薄—中层状钙质岩屑粉砂岩和细粒岩屑石英砂岩互层(图2)[41⁃44]。下伏的天井山组为白云质灰岩或白云岩,顶部为叠层石云质灰岩或白云岩,其沉积时代目前还存在争议,有研究表明,天井山组近顶部叠层石灰岩仍属于卡尼期[21⁃22,29]。此次研究剖面马鞍塘组与天井山组呈整合接触,而在天井山组顶部叠层石云质灰岩中存在一个平行不整合界面,该界面是由于龙门山前陆盆地前缘隆起而导致,标志着研究区由被动大陆边缘向前陆盆地转换[21⁃22]。卡尼期早期(Julian)地层的沉积缺失,使得研究剖面仅发育晚卡尼期(Tuvalian)沉积物[40]。

Figure 2. Late Triassic Carnian stratigraphic column of the Guanyinya section in Hanwang, Mianzhu
-
对绵竹汉旺观音崖剖面开展系统的野外观察、描述,共采集58件岩样,分别制作大薄片(76×50 mm)21件、标准薄片(76×25 mm)35件和抛光面6件。首先利用奥浦偏光显微镜对薄片进行观察、分析和拍照;然后用EPSON GT X980高清扫描仪(3 200 dpi)将薄片扫描成像,导入ImageJ软件统计薄片中碳酸盐颗粒及生物碎屑颗粒粒径和形状。在针对碳酸盐颗粒粒径的镜下随机统计中,观察者可能选择性忽略部分颗粒(例如粒径较小的颗粒)使得统计结果产生偏差,因此,本次使用的颗粒粒径统计方法为:(1)在薄片中选取一定的区域,并统计区域内可见的所有颗粒;(2)使用面积测量法,即在ImageJ中勾画每个颗粒的外轮廓并读取其面积,每张薄片测量400个以上的颗粒,针对位于所选区域边缘的颗粒,区域内面积小于该颗粒二分之一的均不予统计;(3)在ImageJ中识别并提取不同颗粒的轮廓后,测量其Feret’s diameter(费雷特直径,颗粒轮廓上最远两点之间的距离)。测量值统一按照Krumbein[45]提出的计算方法将测量粒径D换算成Φ值(Φ=-log2D),但由于测量的颗粒数据为薄片的截面积,与实际的颗粒大小存在一定的误差,所以运用Friedman[46]提出的公式(Φ=0.381 5+0.902 7×Φ薄片)对数据进行校正,并运用Middleton[47]和Folk et al.[48]的方法分别计算平均粒径、分选、偏度及峰度,最后将ImageJ计算得出的数据circularity(形状系数)、roundness(圆度)、solidity(凸度)和aspect ratio(扁平度)(计算公式如下所示)绘制散点图,用于评估当时水动力条件及不同颗粒的搬运方式[49]。
Circularity=(4π×s)/c²
Roundness=(4×s)/(π×lmax²)
Solidity=s/s拟合椭圆
Aspect ratio=lmax/lmin
式中:s表示颗粒横截面面积;c表示颗粒周长;lmax表示拟合椭圆长轴长;lmin表示拟合椭圆短轴长;s拟合椭圆表示拟合椭圆的面积。
碳酸盐岩薄片的微相描述内容主要包括颗粒类型、基质类型、颗粒含量百分比、颗粒粒径及生物化石特征等微相划分标志,并参考修订后的Wilson标准微相类型将观音崖剖面马鞍塘组碳酸盐岩划分为11种微相类型(MF)[50⁃51]。
-
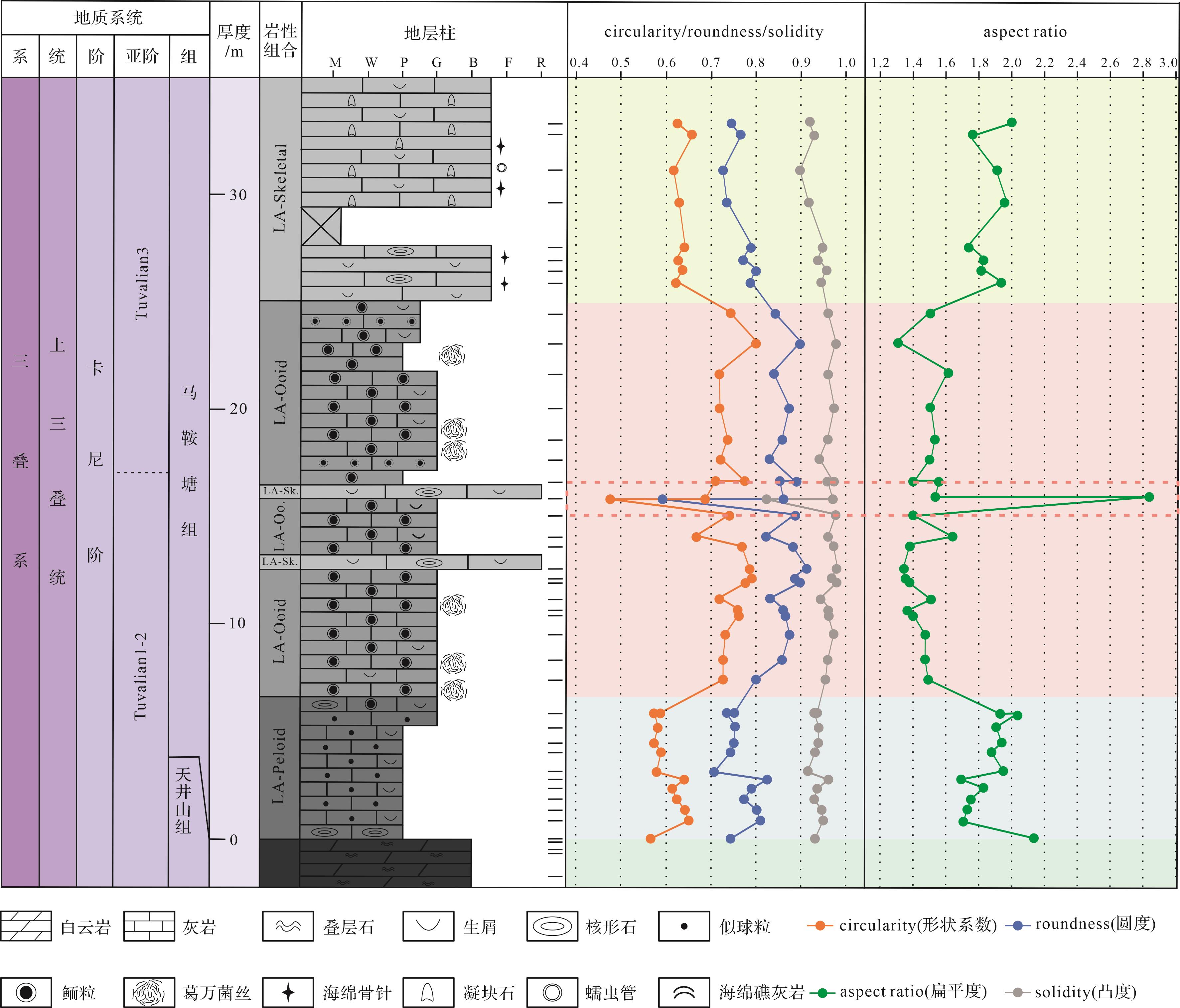
研究区马鞍塘组沉积特征分异明显,结合野外剖面观察,基于室内镜下鉴定结果,识别出11个微相类型(表1),分别是:含包粒似球粒泥粒灰岩—颗粒灰岩(MF1)、含生屑似球粒泥粒灰岩(MF2)、含鲕粒似球粒泥粒灰岩—颗粒灰岩(MF3)、含生屑同心放射鲕泥粒灰岩—颗粒灰岩(MF4)、含生屑表鲕泥粒灰岩—颗粒灰岩(MF5)、含生屑脑状鲕泥粒灰岩(MF6)、含包粒生屑砾状灰岩(MF7)、含生屑包粒泥粒灰岩—漂砾灰岩(MF8)、含生屑凝块石泥粒灰岩—漂砾灰岩(MF9)、海绵微生物黏结灰岩(MF10)、蠕虫管黏结灰岩(MF11)。根据微相特征和相关性,11个微相类型被进一步分为5个岩相组合(Lithofacies Association,LA),分别为似球粒灰岩组合(LA-peloid)、鲕粒灰岩组合(LA-ooid)、生屑灰岩组合(LA-skeletal)、海绵礁组合(LA-sponge reef)以及页岩组合(LA-shale)。
编号 名称 主要特征 沉积相 MF1 含包粒似球粒泥粒 灰岩—颗粒灰岩 颗粒支撑,颗粒间灰泥充填为主,局部有亮晶胶结。包粒粒径主要集中在2 mm左右,核心多为生屑或泥晶,结构复杂,粒间距较大,充填大量粒径0.1~0.2 mm的似球粒(>30%)和部分生物碎屑(15%)。生物碎屑有有孔虫(主要)、腕足、介形虫、海百合茎等,颗粒多破碎,泥晶化,颗粒形态模糊 内缓坡似球粒滩 MF2 含生屑似球粒 泥粒灰岩 颗粒支撑,颗粒间灰泥充填。以似球粒为主,粒径0.1~0.2 mm,存在大量生物碎屑(25%),如有孔虫(主要)、腕足、介形虫、海百合茎等,生屑多破碎,粒径与似球粒相似,大型化石壳体上可见生物钻孔。方解石脉发育 内缓坡似球粒滩 MF3 含鲕粒似球粒泥粒 灰岩—颗粒灰岩 颗粒支撑,颗粒间主要为亮晶胶结,充填少量灰泥。鲕粒粒径极细且均匀,大部分为0.3~0.6 mm,形状不规则,粒径稍大的鲕粒(0.5~0.6 mm)主要发育同心放射状圈层,鲕粒核心多见生屑(棘屑、有孔虫)或灰泥。鲕粒粒间距较大,夹杂粒径较小的碳酸盐颗粒,粒径约0.1 mm,主要为有孔虫、小鲕粒、似球粒及其他细碎生屑,约占整体的10%,未见大粒径生屑 内缓坡似球粒滩 MF4 含生屑同心放射鲕泥粒灰岩—颗粒灰岩 鲕粒圈层以同心放射状为主,直径0.6~1.5 mm,大小混杂,分选较差,小鲕粒(0.6 mm左右)呈团块状集中分布。约有1/4的鲕粒以生屑为核心,其余鲕粒未见核心或以泥晶为核心。生物碎屑较多,主要为棘屑、腹足、腕足、有孔虫、介形虫 内缓坡鲕粒滩 MF5 含生屑表鲕泥粒灰岩—颗粒灰岩 鲕粒以表鲕为主,约占整体的30%,磨圆较好,粒径多介于0.3~0.6 mm。鲕粒核心多为胶结物或者生物碎屑,约占整体的2/3,核心直径明显大于鲕粒包壳的厚度,受核心影响导致鲕粒形状不规则。偶见粒径较大的大鲕粒,粒径接近1 mm,但鲕粒圈层只有0.05 mm左右。生物碎屑丰富,主要生物类型有海百合、藻屑、棘皮、有孔虫等,这些生屑或作为表鲕核心出现,或在鲕粒的间隙单独出现,除藻屑多因重结晶看不清原本结构外,其他生屑大多保存完好 内缓坡鲕粒滩 MF6 含生屑脑状 鲕泥粒灰岩 脑状鲕直径介于0.6~2 mm,鲕粒形状规则,但边缘不光滑,可见部分有孔虫呈包壳状出现在鲕粒内部或边缘;微生物也多参与鲕粒的形成,出现在鲕粒圈层间或鲕粒最外圈层,菌丝保存较好,丝状,高度缠绕。鲕粒粒间夹约10%的小鲕粒、球粒及生屑,粒径介于0.1~0.2 mm。似球粒状泥晶灰岩常见,呈团块状集中分布。生屑多出现在脑状鲕核心或分布在鲕粒粒间,主要有有孔虫、海百合、海胆刺、腕足、藻屑等,鲕粒的形状受核心的影响从而形态不一 内缓坡鲕粒滩 MF7 含包粒生屑砾状灰岩 颗粒支撑,颗粒间为灰泥充填。非骨屑颗粒以包粒为主,大小均匀,多大于2 mm,磨圆较好,多呈球状、椭球状。生物碎屑丰富,含量可达30%,颗粒较大,腕足壳较常见,保存完整,可达1 cm;还出现粒径较大的藻屑,最大可达2 cm,另外可见脑纹状海绵及层孔虫 中缓坡生屑滩 MF8 含生屑包粒泥粒灰岩—漂砾灰岩 主要颗粒为包粒,粒径0.5~3 mm不等,圈层不明显,颜色发黑,形状不规则。包粒间和包粒核心可见生物碎屑,含量约10%,主要有介形虫、海百合、海胆刺、有孔虫、腕足、腹足等,另外海绵骨针在其中杂乱分布 中缓坡生屑滩 MF9 含生屑凝块石泥粒灰岩—漂砾灰岩 靠近礁的部分,出现大量凝块石颗粒,约占整体的10%~20%,粒径为0.2~1.8 mm不等,凝块石为内含细碎生物碎屑的黑色泥晶团块,磨圆中等。在粒间隙常见大量生物碎屑,主要为棘皮类、腹足类、介形类、双壳类、海绵(多为海绵骨针,偶见海绵)等,生物化石破碎,少数保存完好,另外还有大量蠕虫管集中出现,外层厚泥晶套包裹,核心通常充填有亮晶胶结物。似球粒状泥晶大量出现,多呈团块状集中分布 中缓坡生屑滩 MF10 海绵微生物黏结灰岩 基质主要为灰泥,含量可达到70%以上。镜下可见保存较好的海绵及海绵骨针,其他的生物碎屑较少见,偶见保存较好的海百合茎、介形虫、腕足等 外缓坡生物礁 MF11 蠕虫管黏结灰岩 镜下可见蠕虫管局部富集,周围分布有大量似球粒状灰泥。生物碎屑少见,主要为海绵骨针或海绵,偶见海百合茎及腕足壳体的碎片 外缓坡生物礁 Table 1. Characteristics of microfacies from the Ma’antang Formation at the Guanyinya section in Hanwang, Mianzhu
1) 似球粒灰岩组合(LA-peloid)
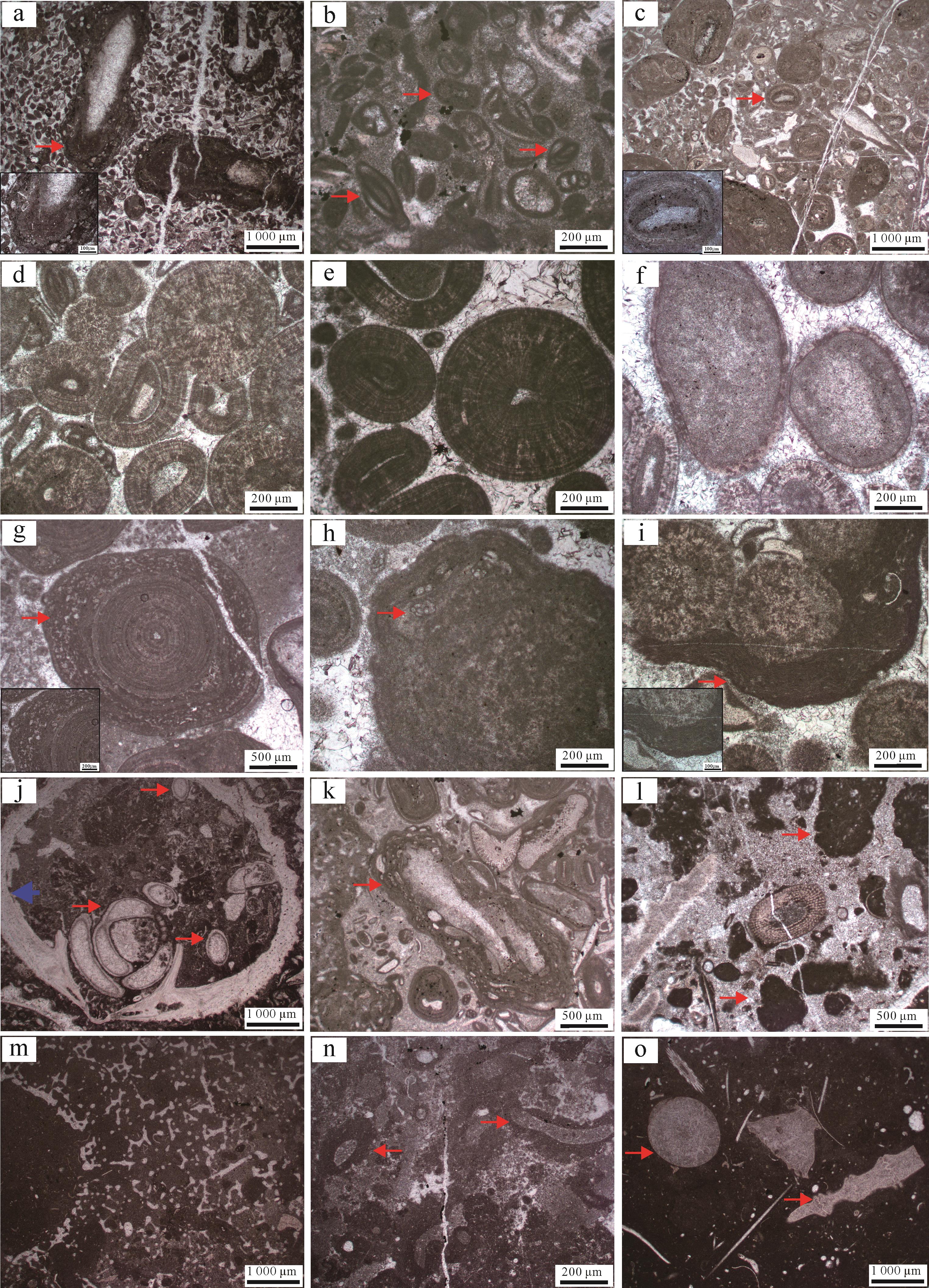
该组合是以似球粒为主的深灰色薄—中层泥粒灰岩—颗粒灰岩,主要颗粒为似球粒,约占整体的30%,其形态及特征与巴哈马似球粒相似,为磨圆和分选都较好的泥晶化细粒暗色团块,粒径约0.2 mm。根据次要颗粒的类型可将该组合分为三个微相:含包粒似球粒泥粒灰岩—颗粒灰岩(MF1)、含生屑似球粒泥粒灰岩(MF2)、含鲕粒似球粒泥粒灰岩—颗粒灰岩(MF3)。在似球粒灰岩组合中,MF1中的次要颗粒是粒径较大、暗色、不规则团块状的包粒(图3a),分选和磨圆都较差,包粒内部未见纹层等结构,但可见包裹其中的有孔虫和海百合茎等生屑;MF2中的次要颗粒是生物碎屑,粒径与似球粒大小相似,约占整体的25%,有孔虫在生屑中占比较高,约占生屑总量的40%,结构保存较好,腔室等细微结构在镜下清晰可见,有孔虫以小栗虫为主(图3b),呈纺锤状;MF3中的次要颗粒为显微结构保存较差的鲕粒,具有丝状同心圈层(图3c)。

Figure 3. Microscopic photographs of different grains
3.1. 绵竹汉旺观音崖剖面马鞍塘组岩石地层特征
-
本组合主要为深灰色中—厚层夹薄层鲕粒粒泥灰岩—颗粒灰岩,根据鲕粒的主要类型可划分为三个微相,分别为含生屑同心放射鲕泥粒灰岩—颗粒灰岩(MF4)、含生屑表鲕泥粒灰岩—颗粒灰岩(MF5)和含生屑脑状鲕泥粒灰岩(MF6)。鲕粒类型丰富,镜下可分辨出五种类型的鲕粒,分别为同心放射鲕、表鲕、脑状鲕、泥晶鲕和复鲕。MF4中主要为同心放射鲕,粒径0.6~1.5 mm,其纹层特征可分为两种:第一种为同心圈层不明显,放射圈层明显(图3d);第二种同心圈层明显,可见到明暗相间的同心条带(图3e)。MF5中以表鲕为主,其特征为核心占整体的比例较大,主要为亮晶方解石或者生物碎屑,外圈层仅占鲕粒半径的1/5~1/3,鲕粒粒径介于0.2~1 mm,圈层无纹层结构,仅见亮色环边,在镜下偶见同心状或同心放射状极薄外圈层(图3f)。MF6以脑状鲕为主,粒径在0.6~2 mm,根据其脑状纹层的特征可分为两种:一种外圈层为同心纹层,纹层弯曲,边缘不光滑,镜下鲕粒边缘具有港湾状、瘤状特征,未见生物参与形成纹层或包壳;另一种脑状鲕的外边缘可见葛万菌缠绕包裹鲕粒,也可见鲕粒周期性地被固着有孔虫结壳包裹而形成不规则边缘(图3g,h)。此外,鲕粒灰岩组合中还常见复鲕和泥晶鲕,复鲕核心为2~3个放射鲕或同心放射鲕粒,整体为粒径较大(>1 mm)的暗色团块,且外圈层形状不规则,镜下可见保存较好的葛万菌(图3i);泥晶鲕通常粒径较小,为0.1~0.5 mm,内部圈层结构不可见,但形状规则,磨圆较好,常与其他类型鲕粒同时出现,镜下常见同心放射鲕与泥晶鲕共存。
-
根据主要颗粒和次要颗粒的不同可分划分为三个微相,分别为含包粒生屑砾状灰岩(MF7)、含生屑包粒泥粒灰岩—漂砾灰岩(MF8)以及含生屑凝块石泥粒灰岩—漂砾灰岩(MF9)。在该组合中颗粒类型以生物碎屑为主,岩石风化面上可见双壳、腕足、层孔虫及海绵等,常与层面近平行排列;镜下可见多种生物碎屑,主要生物类型包括有孔虫、棘皮、腕足、苔藓虫及藻屑等(图3j),生物碎屑占比为10%~20%,有孔虫粒径较小,为0.1~0.2 mm,保存完好;棘皮类碎屑多为海百合茎,粒径分布范围较大,0.2~3.5 mm不等,海胆刺较少见,但保存较完整,在镜下可清晰分辨其脊刺,大小为0.5 mm;苔藓虫保存较好,为多个管状苔藓动物的虫室聚合形成,单个虫室直径为0.05~0.10 mm,聚合体粒径为1.5~2.0 mm;腕足藻屑多保存不完整,粒径不一,最大可达1 cm。此外,在生屑灰岩组合中还存在超过30%的非骨屑颗粒,如包粒和凝块石,包粒粒径0.5~3.0 mm不等,镜下颜色较颗粒间充填的灰泥更暗且形状不规则,常见生屑核心(多为腕足碎片)及有孔虫、葛万菌形成的包壳(图3k);凝块石的特征为黑色泥晶团块,粒径差异较大,从毫米级到厘米级均有,磨圆较差,无核心及圈层结构,团块内含细碎生物碎屑,泥晶化严重(图3l)。另外在生屑灰岩组合中可见保存良好的葛万菌菌丝,多形成颗粒的包壳,或出现在包粒内部,也偶见葛万菌单独出现。
MF7的主要颗粒为保存较好的苔藓虫、藻屑及腕足等,粒径较大,可达厘米级,次要颗粒为包粒,粒径较大,粒径大于2 mm的颗粒超过10%且为颗粒支撑,故命名为含包粒的生屑砾状灰岩;MF8的主要颗粒为包粒,次要颗粒为生物碎屑,主要有藻屑、腕足及海百合茎等,颗粒粒径大于2 mm的超过10%且部分为基质支撑,故命名为含生屑的包粒泥粒灰岩—漂砾灰岩(MF8);MF9中的主要颗粒为凝块石,次要颗粒为生物碎屑,常见海绵骨针和海百合茎。
-
海绵礁灰岩组合中包括两个微相,深灰色厚层海绵微生物黏结灰岩(MF10)和深灰色厚层蠕虫管黏结灰岩(MF11)。该灰岩组合颗粒之间的填隙物主要为灰泥,在镜下可见样品薄片中零散分布的海绵骨针或骨针格架,最典型的骨针格架呈方格网状,是六射海绵的特征,散落的海绵骨针多呈“十字”状,具有四个尖端,长轴长度为0.5~1.0 mm,镜下偶见长3~5 mm的骨针;也可见保存较完整的海绵,骨针有序排列,多为管状海绵的横截面,直径可达1 cm,可见海绵中间用来排水的空腔(图3m)。海绵中常见具有厚泥晶套的蠕虫管,其管径分布范围较大,多根管体呈簇状产出,产状具有定向性,在镜下可见其横截面,管径0.5~5.0 mm,该类蠕虫管无管壁,而是在虫管之外有很厚的泥晶套,泥晶套的厚度达0.1 mm,泥晶套外侧边缘不规则,内侧光滑,这种蠕虫管多与似球粒泥晶一同出现(图3n);在镜下还见到另一种蠕虫管,管径较小约0.5 mm,未见泥晶套,在镜下可见管壁,管壁较薄,约0.02 mm,微亮晶充填,管内空腔被块状方解石充填(图3j)。海绵礁灰岩组合中主要为海绵骨针或海绵碎片,其他生物碎屑较少,偶见海百合及腕足壳体的碎片(图3o)。
5) 页岩组合(LA-shale)
主要由深灰色页岩、互层的砂岩和粉砂岩,及钙质粉砂岩组成。有机质含量较高,海相生物化石、颗粒较大的黄铁矿结核和植物化石极为常见。
上述组合类型在观音崖剖面马鞍塘组的碳酸盐岩沉积序列中相继出现。似球粒灰岩组合在马鞍塘组的底部出现,颗粒类型以非骨屑颗粒为主,如似球粒、包粒等,似球粒粒径在沉积序列中有先减小后增大的趋势,包粒主要集中出现在似球粒灰岩组合的最下部位置,并在似球粒灰岩组合的顶部同鲕粒一起再次集中出现,在该段的灰岩组合中,岩性由含包粒似球粒泥粒灰岩—颗粒灰岩变为含生屑似球粒泥粒灰岩,最后出现含鲕粒似球粒泥粒灰岩—颗粒灰岩;鲕粒灰岩组合沉积在似球粒灰岩组合的上部,颗粒支撑,颗粒之间以亮晶胶结为主,同心放射鲕、表鲕和脑状鲕在该灰岩组合中依次出现,泥晶鲕分布较广,常与其他类型鲕粒同时出现,在鲕粒灰岩组合的沉积序列中泥晶鲕自下往上数量增加,粒径变大,同时在该段的灰岩组合中,岩性由含生屑同心放射鲕泥粒灰岩—颗粒灰岩逐渐演变为含生屑表鲕泥粒灰岩—颗粒灰岩,最后出现含生屑脑状鲕泥粒灰岩。在鲕粒灰岩组合中夹两层生屑灰岩,厚约0.4 m,野外可见较大的双壳、腕足、层孔虫及海绵等生屑,常与层面近平行排列,镜下还见到棘屑、腹足、腕足、有孔虫、介形虫、海绵、蠕虫管,和个别粒径1~2 cm的苔藓虫、海绵及重结晶的藻屑。生屑灰岩分隔开的鲕粒灰岩中颗粒之间灰泥含量增加,由鲕粒颗粒灰岩变为鲕粒泥粒灰岩;生屑灰岩组合主要沉积在鲕粒灰岩组合之上,生屑灰岩组合中除了出现大量生物碎屑外,还存在大量包粒和凝块石,包粒出现在该灰岩组合的下部,岩性为含生屑包粒泥粒灰岩—漂砾灰岩,往上包粒消失,由凝块石取而代之,岩性转变为含生屑凝块石泥粒灰岩—漂砾灰岩;最后出现的海绵礁灰岩组合发育在生屑灰岩组合之上,并呈透镜状一部分沉积在页岩组合中,海绵礁灰岩组合岩性分界明显,下部为海绵微生物黏结灰岩,上部转变为蠕虫管黏结灰岩。
-
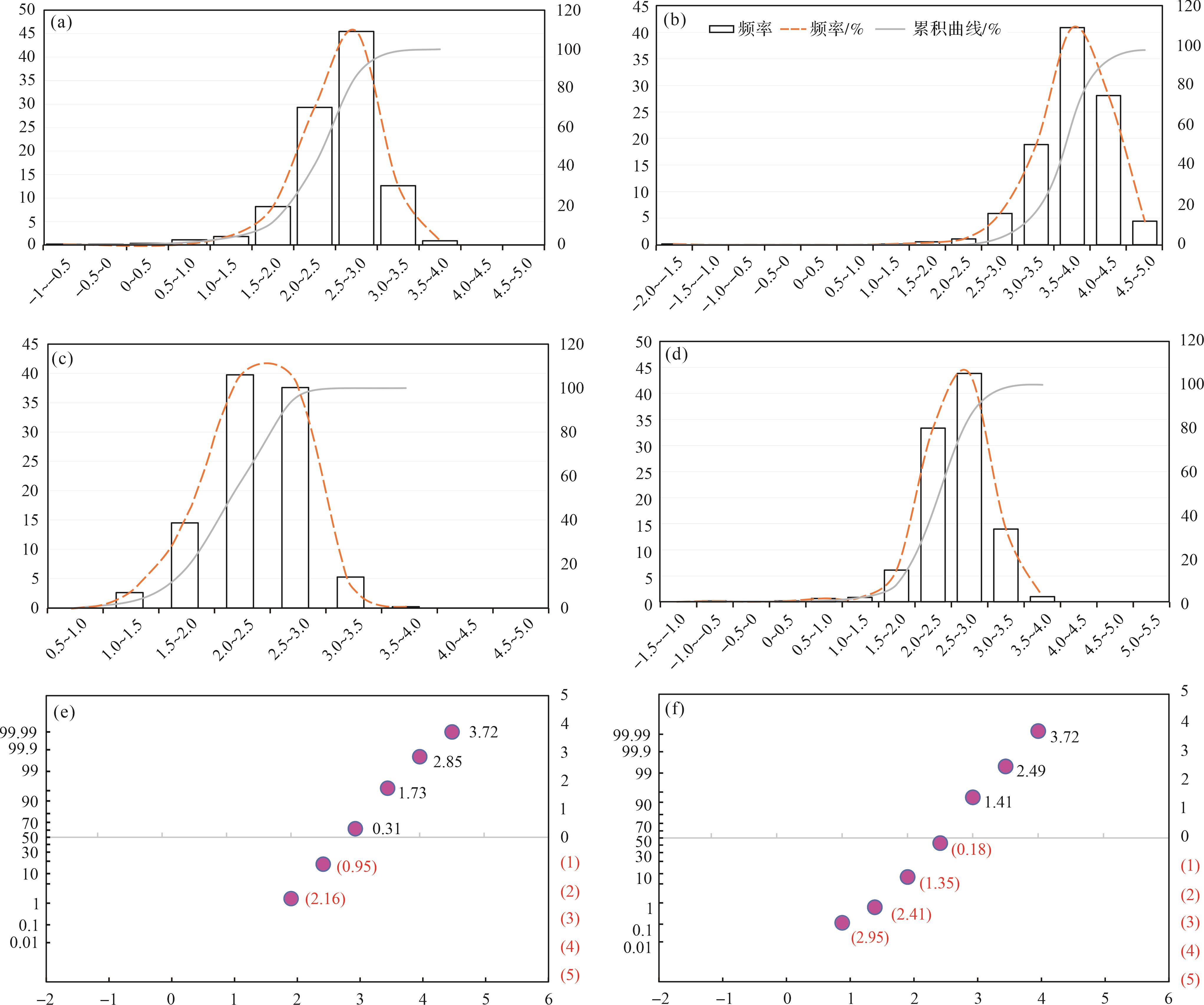
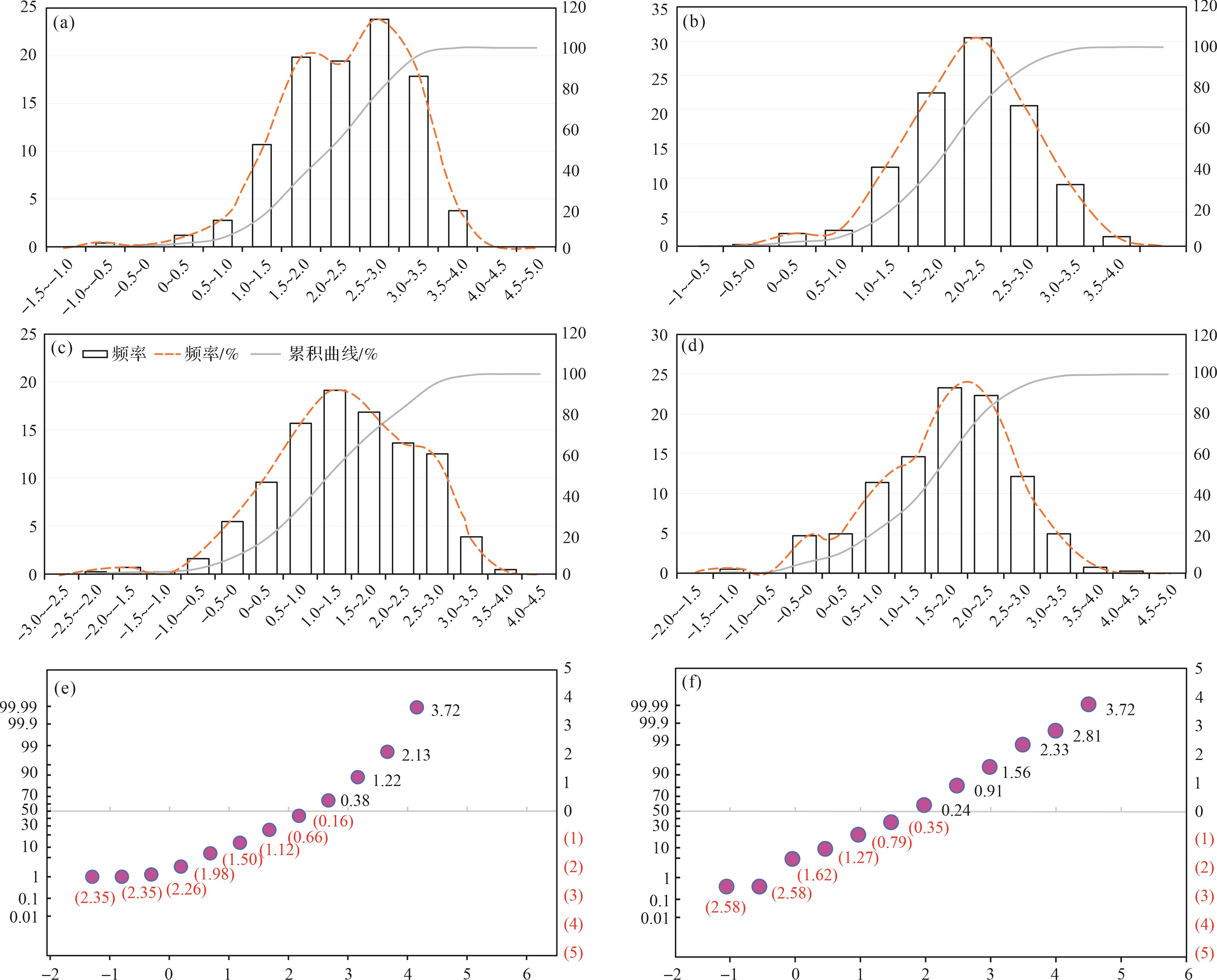
似球粒灰岩组合。镜下粒径统计结果表明,似球粒灰岩组合部分的颗粒粒径主要集中在44~354 μm,在粒度分布直方图及累积曲线中,主峰先往右偏移后往左偏移且逐渐不突出,反映Φ值先增大后减小,颗粒粒径先减小后增大,这与野外特征一致(图4a~d)。峰值由样品GYYD-06中的177~333 μm减小至GYYD-09样品的67~88 μm,最后在GYYD-16-1样品中恢复至177~333 μm(图4a~d)。镜下可观察到GYYD-06和GYYD-16-1样品中出现了粒径较大的包粒、鲕粒及生物碎屑,根据统计结果其粒径最大可达到1 414~2 000 μm(图4a,d)。

Figure 4. Carbonate grain size histogram, cumulative curve, and probability value cumulative curve of peloid lithofacies association
概率值累积曲线特征为粒度较细,Φ值均大于1,GYYD-09到GYYD-14号样品曲线以“一段式”为主,且“一段式”的斜率较大,曲线斜率最高可达60°,反映了似球粒颗粒分选性较好的特点,搬运方式以跳跃搬运为主(图4e,f)。动荡的水体易对跳跃搬运的颗粒进行分选,跳跃搬运的组分往往是沉积物中分选最好的部分。
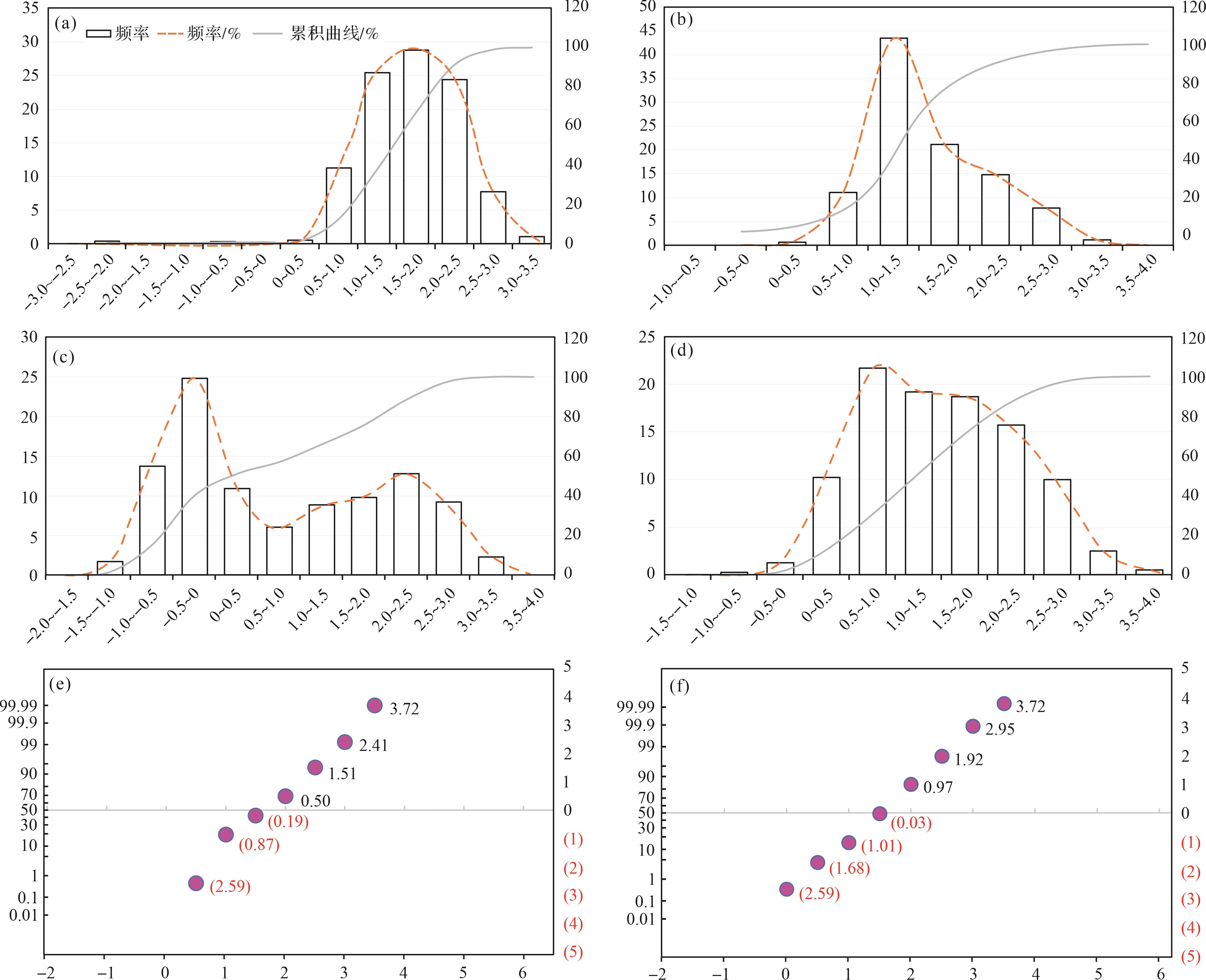
鲕粒灰岩组合。在整个鲕粒灰岩组合的样品中未见鲕粒粒径出现规律性变化。在镜下可见鲕粒分选较差,粒径统计结果和镜下观察到的现象一致,鲕粒的粒度分布范围较大,从88~2 000 μm均有出现(图5a~d)。在粒度直方图中,仅在3~4个样品粒径统计直方图中出现单峰值,单峰值为354~500 μm,其余样品的粒径统计直方图为多峰或峰值不明显,峰值介于177~707 μm,这也符合镜下看到的颗粒分选较差这一特征,GYYD-34号样品中出现大量粒径较大的鲕粒,最大可达2 000 μm(图5c)。其中该灰岩组合中可能偶尔出现保存较完整的生物碎屑,如在GYYD-18号样品中出现的腕足壳体,粒径可达5 000 μm,同时在大粒径鲕粒的粒间夹杂其他碳酸盐颗粒,如GYYD-34号样品中的小粒径鲕粒和生物碎屑等,导致该灰岩组合分选极差,可分出十个粒度区间(图5a,c)。概率值累积曲线特点为Φ值最小为0,该组合颗粒粒径较似球粒灰岩组合部分有所增加,曲线以不标准的“一段式”为主,斜率较小,接近30°,反映了鲕粒灰岩组合中颗粒分选性较差的特点,搬运方式以跳跃搬运为主,几乎未见滚动和悬浮颗粒(图5e,f)。

Figure 5. Carbonate grain size histogram, cumulative curve, and probability value cumulative curve of ooid lithofacies association
生屑灰岩组合。在该灰岩组合中出现的颗粒主要有凝块石、包粒及生物碎屑,样品统计平均值分别为194 μm、274 μm、221 μm、282 μm、239 μm、362 μm和366 μm,整体上有稍微变大的趋势。颗粒粒径分布范围较大,从65~4 000 μm皆有分布,粒度区间多达8~11个,峰值多不明显,未出现双峰或多峰,从粒径统计上分析该段分选极差,存在大粒径的包粒或凝块石,这与镜下观察到的现象一致(图6a~d)。

Figure 6. Carbonate grain size histogram, cumulative curve, and probability value cumulative curve of skeletal lithofacies association
概率值累积曲线特点为曲线以“两段式”为主,曲线斜率较小,反映了该灰岩组合颗粒分选性较差的特点,搬运方式以滚动和跳跃搬运为主,几乎未见悬浮颗粒(图6e,f)。
粒度平均值、分选、偏度及峰度统计结果显示(图7),在似球粒灰岩组合中,含包粒似球粒泥粒灰岩—颗粒灰岩间隔出现,导致样品的平均值、分选、偏度及峰度存在一定幅度的波动。含包粒似球粒泥粒灰岩—颗粒灰岩样品的平均粒径较大,为169~346 μm,含生屑似球粒泥粒灰岩—颗粒灰岩和含鲕粒似球粒泥粒灰岩—颗粒灰岩样品平均粒径值则集中在82~173 μm;似球粒灰岩组合样品分选值基本小于其他灰岩组合样品分选值,反映其分选与其他灰岩组合相比较好;该灰岩组合中含包粒似球粒泥粒灰岩—颗粒灰岩的偏度值主要为正偏态,峰偏向粗粒度一侧,即存在大量包粒,而含生屑似球粒泥粒灰岩—颗粒灰岩和含鲕粒似球粒泥粒灰岩—颗粒灰岩的偏度值小于零,为负偏态,沉积物以细粒(似球粒)为主;似球粒灰岩组合中底部样品峰度值存在小幅度波动,说明是由多种类型的颗粒混合沉积而成。

Figure 7. Characteristics of carbonate grain size distribution
鲕粒灰岩组合的平均值集中在285~351 μm,该组合中的颗粒平均粒径大于似球粒灰岩组合中的颗粒平均粒径,该组合中含生屑同心放射鲕泥粒灰岩—颗粒灰岩和含生屑表鲕泥粒灰岩—颗粒灰岩部分样品分选值变化不大,说明分选较好,含生屑脑状鲕灰岩部分分选值发生波动,分选性变差;偏度值集中在0,说明颗粒粗细均匀。在样品GYYD-35处镜下可观察到鲕粒双峰态明显,小粒径的鲕粒和似球粒局部集中分布在大粒径的鲕粒间隙之间,这一特征同样可以在平均值、分选、偏度统计图中显示出来:平均粒径Φ值稍有减小(实际值增大),分选值达到1.25,分选较差,偏度值大于0,以粗粒径颗粒为主。
生屑灰岩组合的平均值集中在195~608 μm,样品平均粒径最大可达608 μm,明显大于其他两个灰岩组合的颗粒平均粒径。样品分选值大于似球粒灰岩组合以及鲕粒灰岩组合,说明该灰岩组合分选较以上两种更差。
-
定量描述颗粒形态特征的参数形状系数(circularity)、圆度(roundness)、凸度(solidity)和扁平度(aspect ratio)是由ImageJ软件直接计算得出,将其投点得到下图(图8):在图中似球粒灰岩组合中出现包粒部分circularity值为0.5~0.6,roundness为0.7~0.8,aspect ratio为1.5~2.0,颗粒极不规则,磨圆中等,以似球粒为主的颗粒aspect ratio接近2,以形状规则、磨圆较好的扁长形椭圆为主。鲕粒灰岩组合部分的整体碳酸盐颗粒circularity值集中在0.7~0.8,roundness为0.8~0.9,solidity略大于似球粒灰岩组合部分,aspect ratio集中在1.5,说明颗粒磨圆较似球粒灰岩组合部分更好,形状以椭圆为主但比似球粒更饱满、更圆。在生屑灰岩组合部分中,circularity值为0.6~0.7,roundness为0.7~0.8,aspect ratio为1.5~2.0,颗粒极不规则,磨圆较差。其中GYYD-30号样品中出现极不规则生屑,导致四个参数有较大变化,这一特征与镜下观察到的一致(如图8中红色虚线范围)。

Figure 8. Characteristics of carbonate grain size shape
3.2. 颗粒统计特征
3.2.1. 颗粒粒径特征
3.2.2. 颗粒形态特征
-
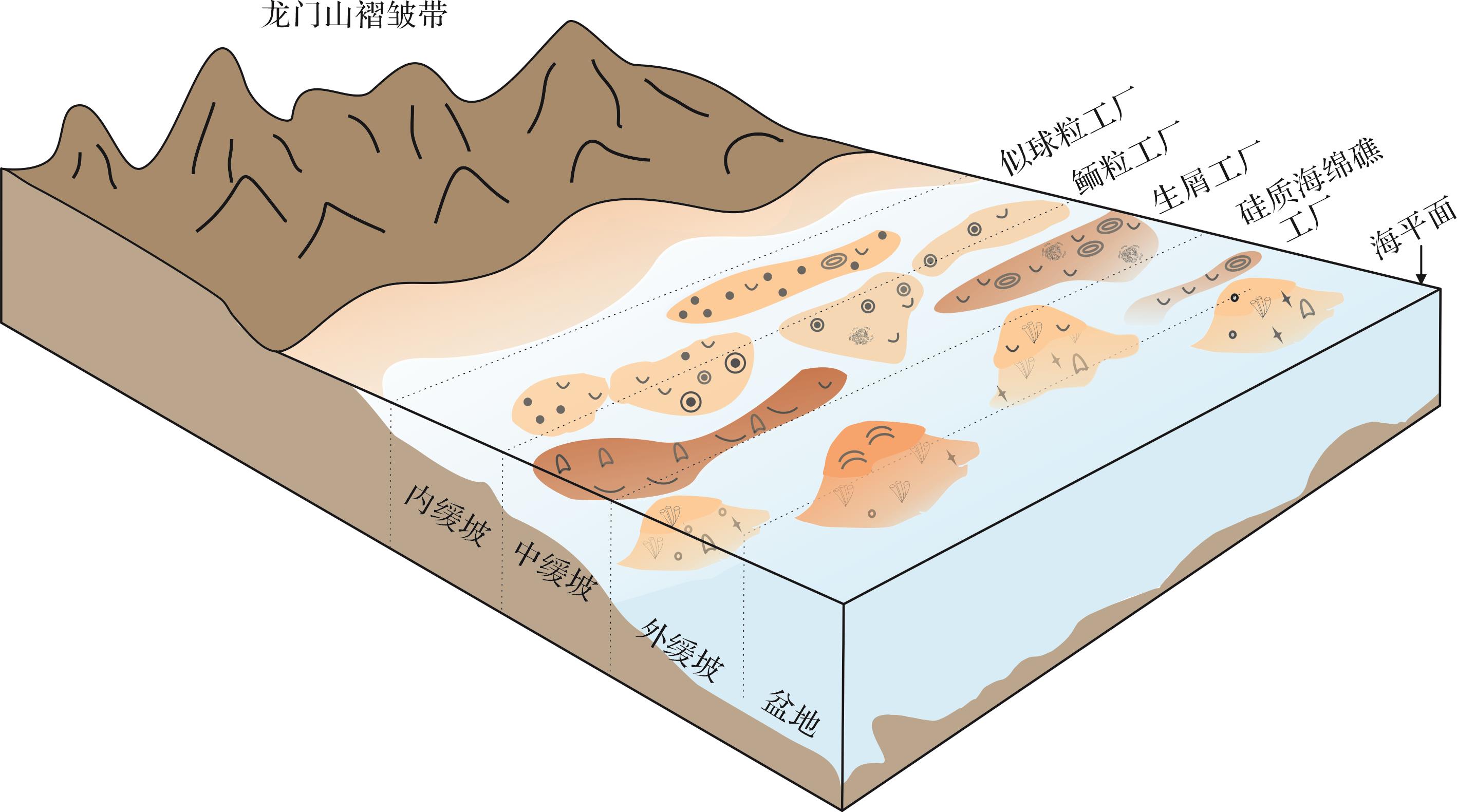
研究区热带浅水碳酸盐生产工厂可进一步划分为以似球粒、鲕粒、生屑及生物礁等颗粒组合表征的特异化工厂类型,指示了不同的碳酸盐生产过程。沉积序列中纵向短时间尺度内多种特异化工厂的交替出现,可以反映多种碳酸盐生产模式在横向空间上的共存及相互转化(图9)。似球粒组合工厂主要颗粒为似球粒,同时存在粒径较大的包粒,生物碎屑以底栖有孔虫为主,可以解释为沉积在中—高能环境,位于靠近潟湖的局限内缓坡的浪基面以上,大量存在的包粒则是由微钻孔生物在极浅的环境中通过泥晶化作用形成的,生物碎屑分选和磨圆都较好,同样也说明了这套沉积物形成于相对高能的环境中[52⁃54];鲕粒组合工厂中的主要颗粒为鲕粒,颗粒间多以亮晶胶结为主,表明此时水体能量达到最高,且长时间维持一种氧分充分、营养水平低但水体能量较强的状态,同时鲕粒的双峰态分布说明了鲕粒组合工厂可能发育在受风暴影响的内缓坡高能浅滩或海岸环境,不一定为原地工厂[55⁃56];生屑组合工厂以颗粒支撑为主,颗粒间多充填灰泥,说明此时的水深大于似球粒组合工厂和鲕粒组合工厂。此外,大量的微生物形成的碳酸盐颗粒以及种类丰富的海洋生物表明了沉积环境为开放的低—中能的内缓坡到中缓坡[57];海绵礁组合工厂主要由蠕虫管黏结灰岩、海绵微生物黏结灰岩组成,沉积环境指示中缓坡的外侧,李勇等[58]根据该区原位海绵礁的厚度,估算了海绵生长的水深在150 m以上。自海绵礁开始生长,细石英粉砂开始出现[23],陆源碎屑物质的出现可能为海绵或其他异养生物提供了营养,也可能正是因为大量陆源碎屑物质的输入导致了海绵礁的消亡[20];随后,在海绵礁上部覆盖的页岩和泥质粉砂岩中发现大量开阔海环境指相化石,如菊石,表明研究区海平面持续上升[24]。

Figure 9. Sedimentary pattern and distribution of subdivided carbonate factories from the Ma’antang Formation in western Sichuan
-
四川盆地西北部的绵竹汉旺地区马鞍塘组沉积的碳酸盐厚度约为70 m[20,25],同沉积期出现的多个细分特异化工厂类型,指示了不同环境下碳酸盐生产方式的差异。
似球粒组合工厂主要建造者包括似球粒、包粒及生屑颗粒,主要通过生物诱导和生物控制矿化共同参与形成。其中似球粒主要是由生屑和鲕粒首先经过藻类钻孔,然后被泥晶方解石充填,导致了显微结构完全消失,产生的圆形泥晶颗粒,为典型的巴哈马似球粒[51,54];包粒是由微生物和其他结壳类生物形成的结核状包壳颗粒,研究区出现的包粒根据形成包壳生物不同可分为两类,即葛万菌包粒和有孔虫包粒,包壳的圈层由固着的生物壳体(即葛万菌和有孔虫)形成,通过黏结或捕获细粒沉积物或促进同沉积期沉积物发生矿化作用[59⁃60];大量包粒的存在和颗粒普遍的泥晶化表明颗粒的长时间静置,使得藻类等钻孔生物有足够的时间对颗粒进行钻孔和磨蚀[54]。
鲕粒组合工厂是研究区的主要细分工厂类型,鲕粒的生长需要合适的碳酸盐颗粒作为核心,碳酸盐晶体围绕核心发生化学沉淀并聚集形成纹层,这一过程要求处于碳酸盐超饱和水体中且处于悬浮搅动阶段[61]。鲕粒的形成通常被认为是一个非生物成因的无机过程,但鲕粒圈层间和外部发育的微生物结构说明存在微生物参与的矿化作用[55];脑纹状鲕粒与纹层状鲕粒内部结构高度相似,外凸的纹层结构可能反映了微生物群落对光照和营养的竞争;复鲕是经过黏结、凝聚和胶结作用等多个阶段最后形成的[62]。
生屑组合工厂中碳酸盐的沉淀与生物控制的骨骼或壳体的钙化作用和微生物诱导矿化有关[8]。骨屑颗粒是通过生物控制矿化作用形成的,在生屑组合工厂中,光自养型生物大量存在,如藻类;另外,还有与藻类共生的动物,包括底栖有孔虫、软体类和部分造礁海绵[10]。凝块石为一种不具纹层构造,形状大小各异,磨圆较差的泥微晶团块,可能是微生物在生命活动中沉淀并黏结灰泥而形成的,或和生物腐烂分解过程中的有机质降解有关[8,63⁃64]。
海绵礁组合工厂中主要的生产者为光自养型生物和后生动物。研究区主要造礁生物除海绵外,与之共生的还有大量的蠕虫管和微生物,蠕虫管和微生物通过捕获细粒物质并诱导碳酸盐沉淀,形成了特有的黏结—障积海绵礁[59,63]。棘皮类、腹足类、介形类、双壳类、海绵(多为海绵骨针,偶见海绵)等的存在说明海绵礁的生长为底栖后生生物提供了适宜的生存环境[39⁃41],也成为海绵礁组合工厂中的一部分碳酸盐颗粒的来源。
-
研究区碳酸盐工厂转变的主要影响因素为海平面的变化,海平面的变化一方面直接影响着水体能量的变化和海洋生物的种类,另一方面调控着碳酸盐生产的环境[65⁃66]。汉旺地区在晚三叠世之前经历了一次海平面下降,而在晚三叠世卡尼期,由于龙门山造山楔构造负载引起扬子板块西部的挠曲沉降,从而导致研究区水深不断加深(图2)[40]。研究区发生了短暂的碳酸盐台地暴露及随后持续的海侵,碳酸盐的生产经历了下降然后恢复的过程[15⁃19]。
海平面变化同时影响着温度和碳酸盐饱和度[61,67]。温度主要以两种方式影响碳酸盐生产者:首先,温度直接影响了生物种类,且变温生物新陈代谢相关的化学反应主要由温度决定[68];此外,温度控制了二氧化碳的溶解度和碳酸钙的饱和度,从而影响海洋钙化和维持碳酸盐骨骼所需要的物质来源和反应能量[61,69⁃70]。研究区的暖水环境适合多种海洋生物生存并进行生物钙化,同时暖水条件下也有利于维持高的碳酸盐饱和度,海水饱和度对不同颗粒的形成起到一定的促进或抑制作用。研究表明,在Tuvalian时期,海水表层温度经历了先上升后下降的过程,海水温度在Tuvalian1到Tuvalian2时期升高,并在Tuvalian3时期开始下降,因此似球粒组合工厂到鲕粒组合工厂的转变被认为可能是由于水温升高引起碳酸盐饱和度升高的结果[21,33,71];随后研究区的海平面加深的同时海水温度持续降低,鲕粒组合工厂的消亡正是水体能量降低以及碳酸盐高饱和状态被破坏的一个实例,水体的加深改变了生物组合,为海绵礁组合工厂的发育创造了条件[22,25,34]。
海平面变化直接控制着水体能量,并通过影响颗粒的大小、分选和磨圆,以及搬运形式等影响碳酸盐工厂的类型和构成,此外,风暴在其中也起到一定的作用,还通过搅动、搬运等形式影响碳酸盐工厂的颗粒组成[72⁃74]。针对水动力的讨论主要为在Tuvalian1时期研究区不同水动力强度对碳酸盐颗粒的影响:似球粒组合工厂中碳酸盐颗粒的平均粒径为65~272 μm,粒径较小,且颗粒的分选最好,aspect ratio接近2,以形状规则、磨圆较好的扁长形椭圆为主,其搬运方式主要为跳跃搬运,说明其发育在中—高能环境;鲕粒组合工厂中的颗粒平均粒径介于285~351 μm,较似球粒组合工厂有所增加,roundness为0.8~0.9,solidity略大于似球粒灰岩组合部分,aspect ratio小于似球粒组合工厂,说明碳酸盐颗粒磨圆较似球粒灰岩组合部分更好,形状更加饱满;搬运形式一般为滚动或跳跃搬运,在其发育时期,水动力强度应大于似球粒组合工厂。在鲕粒组合工厂中鲕粒粒径出现双峰态分布,小粒径的鲕粒和似球粒局部集中分布在大粒径的鲕粒间隙之间,分选值达到1.25,偏度值大于0,以粒径较大的鲕粒为主,另外还出现多个的生屑富集带,镜下生屑粒径最大可达7 mm,野外可见厘米级的海绵及层孔虫等造礁生物碎屑,以上现象说明在鲕粒工厂发育的过程中疑似受到风暴作用的影响[20,39],并且可能在横向上与生物礁共存;生屑组合工厂中出现的碳酸盐颗粒除生物碎屑外,还存在大量的包粒和凝块石,平均粒径集中在195~608 μm,明显大于其他两种组合工厂的颗粒平均粒径,circularity值为0.6~0.7,roundness为0.7~0.8,aspect ratio为1.5~2,指示颗粒极不规则,磨圆较差,再加上部分颗粒间由灰泥充填,代表着水深的加深,呈现一种水动力较弱的状态。
而各个细分工厂在沉积序列纵向上的快速变化也可以指示它们在沉积体系中横向上的共生关系。因此,较短时间尺度下细分工厂类型的转变可能主要与区域海平面变化控制的横向生产工厂类型的迁移有关,而与极端事件等关系不大。
-
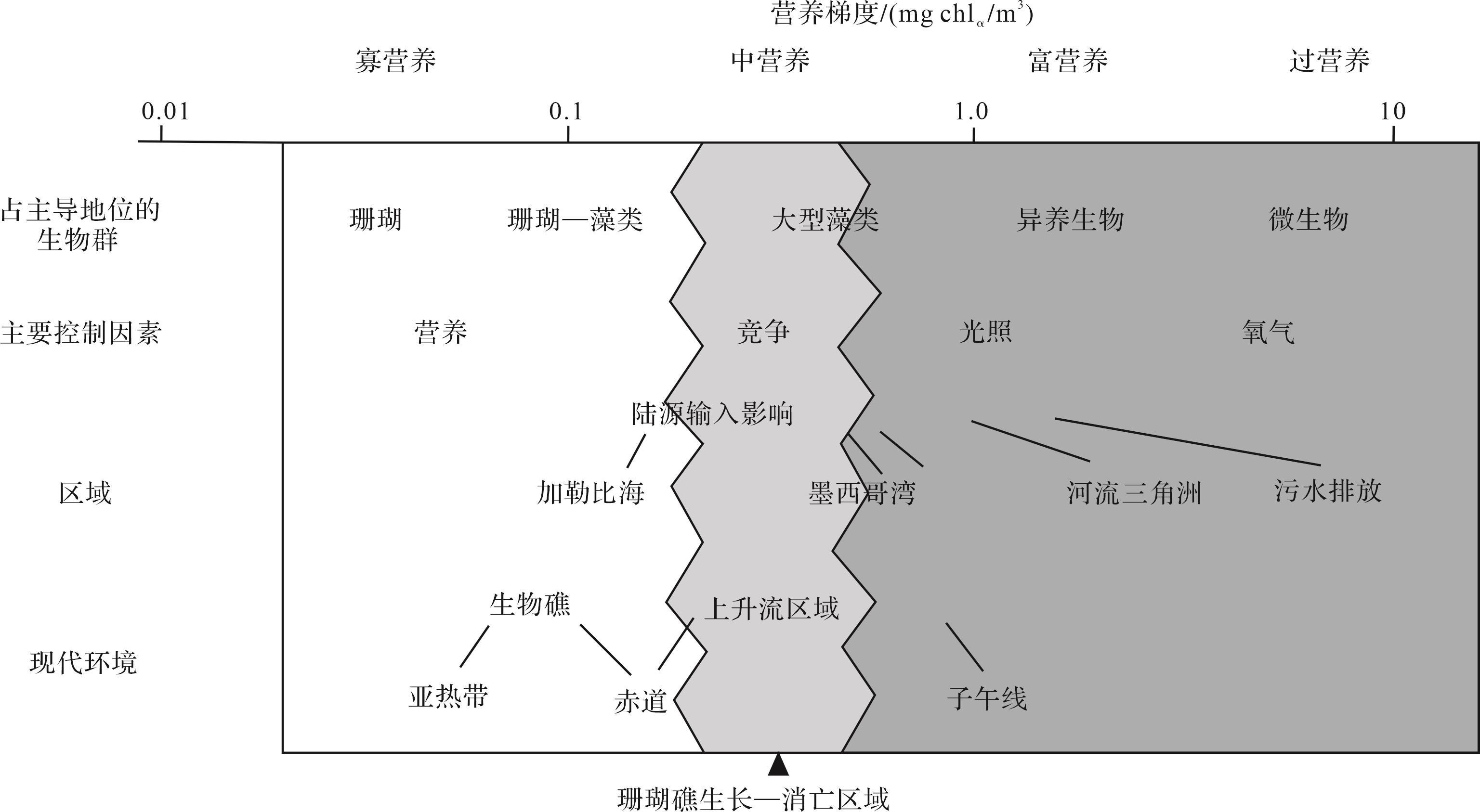
营养水平是调节碳酸盐生产的关键因素,控制了浅水环境中不同工厂类型的形成[75]。“营养物质”可以用来描述环境中的有机物,也可以包括通过光能和化学能合成有机物的无机营养物质,其主要来源为陆源物质或有机质的输入,从而影响底栖生物群的多样性、优势性和稳定性[10]。碳酸盐岩和生物礁的发育往往受营养梯度的影响,从生态学的角度来看,热带水体中群落结构的主要转变都是随着环境中营养梯度的变化而改变,从寡营养到中营养,从中营养到富营养,从富营养到过营养(图10)[76]。在这四种梯度中,每一种都具有不同的限制因素,例如在寡营养区域,几乎不存在营养资源的竞争,营养资源的竞争主要发生在中营养区域,而富营养区域由于浮游生物的大量存在,导致底栖生物极大地受到光照的限制。过营养环境的地表水中有机碳过饱和,底栖生物的生存受到氧气含量的影响[77⁃78]。球粒、包粒的出现往往伴随着高营养水平,而营养水平低就会出现亮晶胶结的鲕粒和生物礁的发育,而微生物主导的礁往往形成于富营养环境,海绵为主导的生物礁可能形成于中等营养水平[79⁃81]。因此研究区在卡尼期可以认为是一个从相对中等营养过渡到低营养再到中高营养水平的过程。研究区的似球粒组合工厂的发育过程往往需要一定的营养水平,部分营养物质的输入,极大地促进了钻孔生物的繁盛,球粒越致密,就是泥晶化越强的表现,同时这也是一些微生物适应强水动力的指示[23,40]。在鲕粒组合工厂部分,频繁出现的风暴不仅造成了鲕粒的双峰态分布,也带来了大量的生物碎屑,同时也为养分含量相对较低的鲕粒滩带来一定的营养盐,因此能够偶尔看到微生物在鲕粒组合工厂中局部集中出现[20,75]。陆源碎屑含量在生屑组合工厂及海绵礁组合工厂的发育期间持续增加,为其带来大量的营养物质,因此,以上两种细分工厂可能发育在中—高营养环境中。同时,还可能存在由于有机质降解(如生物礁中的海绵死亡后)导致局部环境营养水平升高的情况,这一点也可以用来解释海绵微生物黏结灰岩中大量微生物相关的灰泥组分以及富有机质的凝块石存在的现象。

Figure 10. Nutrient gradients in low latitude waters (modified from reference [76])
4.1. 川西北汉旺观音崖剖面马鞍塘组浅水碳酸盐工厂的构成
4.2. 同沉积期不同环境下碳酸盐生产方式的差异
4.3. 碳酸盐生产工厂的主要影响因素
4.3.1. 海平面变化及相关因素
4.3.2. 营养水平的变化
-
(1) 四川盆地西北部的绵竹汉旺地区马鞍塘组碳酸盐岩沉积可识别出11个碳酸盐微相,根据微相特征和相关性,可进一步归为5类岩相组合,分别为似球粒灰岩组合、鲕粒灰岩组合、生屑灰岩组合、海绵礁组合及页岩组合。
(2) 研究区以热带浅水碳酸盐岩沉积为主,可进一步划分为以似球粒、鲕粒、生屑及生物礁等特异化工厂类型,能够指示不同的碳酸盐生产过程。其中似球粒组合工厂发育在位于靠近潟湖的局限内缓坡浪基面以上的中—高能环境,其形成多与微生物钻孔、包壳以及黏结作用有关;鲕粒生长在高碳酸盐饱和度环境中且处于悬浮搅动阶段,因此鲕粒组合工厂可能发育在受风暴影响的内缓坡高能浅滩或海岸环境;生屑组合工厂发育在开放的低—中能的内缓坡到中缓坡环境,其形成往往与生物诱导和生物控制的矿化作用有关;海绵礁组合工厂发育在中缓坡外侧的低能环境中。
(3) 研究区热带浅水碳酸盐工厂发育及转换更多地受区域海平面变化控制。频繁的海平面变化导致碳酸盐矿物饱和度、水体能量等因素快速变化,进而驱动了具体细分工厂的转变。另外,营养水平是调节碳酸盐生产的关键因素,可能控制了浅水环境中不同工厂类型的形成。研究区沉积序列上众多细分工厂的快速变化指示了其在横向上的共生关系。因此,细分工厂类型在短时间内的转变可能主要由区域海平面变化控制的横向生产工厂类型的迁移引起,而与极端事件等无关。

 DownLoad:
DownLoad: