



HTML
-
距今约201 Ma的三叠纪—侏罗纪之交(Triassic-Jurassic boundary, TJB)发生了显生宙以来五次最大规模之一的生物集群灭绝事件,在属一级别上至少有50%的海洋生物和陆地生物灭绝[1⁃2],被称为三叠纪末期生物大灭绝(end-Triassic mass extinction, ETE)[3⁃6]。同期,中大西洋火成岩省(Central Atlantic Magmatic Province, CAMP)爆发和泛大陆(Pangea)进一步裂解致使中大西洋开启[7],深刻地影响了其后约2亿年全球板块构造格局。根据植物叶片化石的外表皮细胞上的气孔密度(stomatal density, SD)和气孔指数(stomatal index, SI),推断早侏罗世初大气中的CO2浓度升高到了晚三叠世末的2~4倍,浓度高达2 500×10-6以上[8⁃11]。根据升高的CO2浓度,推测对应的全球平均温度升高3 ℃~4 ℃[8]。在随着泛大陆裂解的构造作用和温室效应主导的气候共同作用下,早侏罗世全球海平面总体呈上升趋势,但有相对短时的海平面下降[12⁃13];有研究认为海平面变化引起大洋缺氧,对ETE起到了关键作用[14⁃17],但也有学者报道了不同的证据和推论[18]。同期,无论热带和非热带的海洋中碳酸盐岩相或硅质岩相环境中,海绵类、双壳类、珊瑚、腕足类都发生显著的灭绝[19];海洋中的初级生产者,由钙质超微类转换成有机质壳类的鞭毛藻和疑源类为主[20],暗示海洋酸化的发生。陆地上出现与白垩纪末蕨类植物爆发类似的现象,有些地区记录到短暂的蕨类植物孢粉含量激增(持续小于400 kyr)[21⁃23]。由地层中丝炭含量对岩石质量做归一化的统计数据显示,森林火灾发生的频率显著增加[24]。地球表层系统可交换的无机碳库和有机碳库中碳同位素强烈波动(carbon isotope excursion, CIE)[4,15,18,25⁃29],显著波动的CIE与ETE和TJB密切关联[4,30⁃36],并在全球范围内被广泛地记录[4,8,15,37]。具体地,在地层中从下至上包括初始负偏(I-CIE)和主负偏(M-CIE)两次,它们可与CAMP三个主要的喷发期次中两次相相关;I-CIE可能对应CAMP最初的侵位和喷发,M-ICE可能对应CAMP喷发最剧烈期次之后(第三期之前)的生态系统开始恢复时期[38⁃43]。
目前,在奥地利Kuhjoch的三叠系—侏罗系GSSP(“金钉子”)剖面和同盆地内邻近剖面,取得了若干条厘米级分辨率有机碳同位素地层曲线[35]。另外,基于C3植物的有机碳同位素地层学取得了多方面进展,包括:植物从大气固碳过程中的同位素分馏的植物生理学研究[44⁃45]、正演的植物培育实验[46⁃47]、微生物作用和成岩作用过程对于样品记录原始值改变的影响[48⁃51]以及地质实证数据[52⁃54]和地质实证数据的回归分析[55]方面的成果,且大气均一周期远小于百年尺度[56]。以上成果为利用有机碳同位素地层学恢复地史时期大气中CO2的δ13C演化以及进行全球等时地层对比夯实了基础。从地球系统科学的角度对重大地质事件深入的、全球性的研究,依赖于不同剖面之间的精细等时对比,进而区分不同剖面记录的地质事件的先后次序,推断其因果关系和第一驱动力。但通常由于缺乏跨海陆的、全球性可对比的化石,使得海陆间地层对比存在巨大挑战。在中国晚三叠世末—早侏罗世主要发育陆相沉积,生物地层很难与TJB的GSSP海相地层有效地对比[57]。本文以新疆塔里木盆地库车河剖面为例,尝试利用陆相有机碳同位素地层学进行跨海陆地层的精细对比。
-
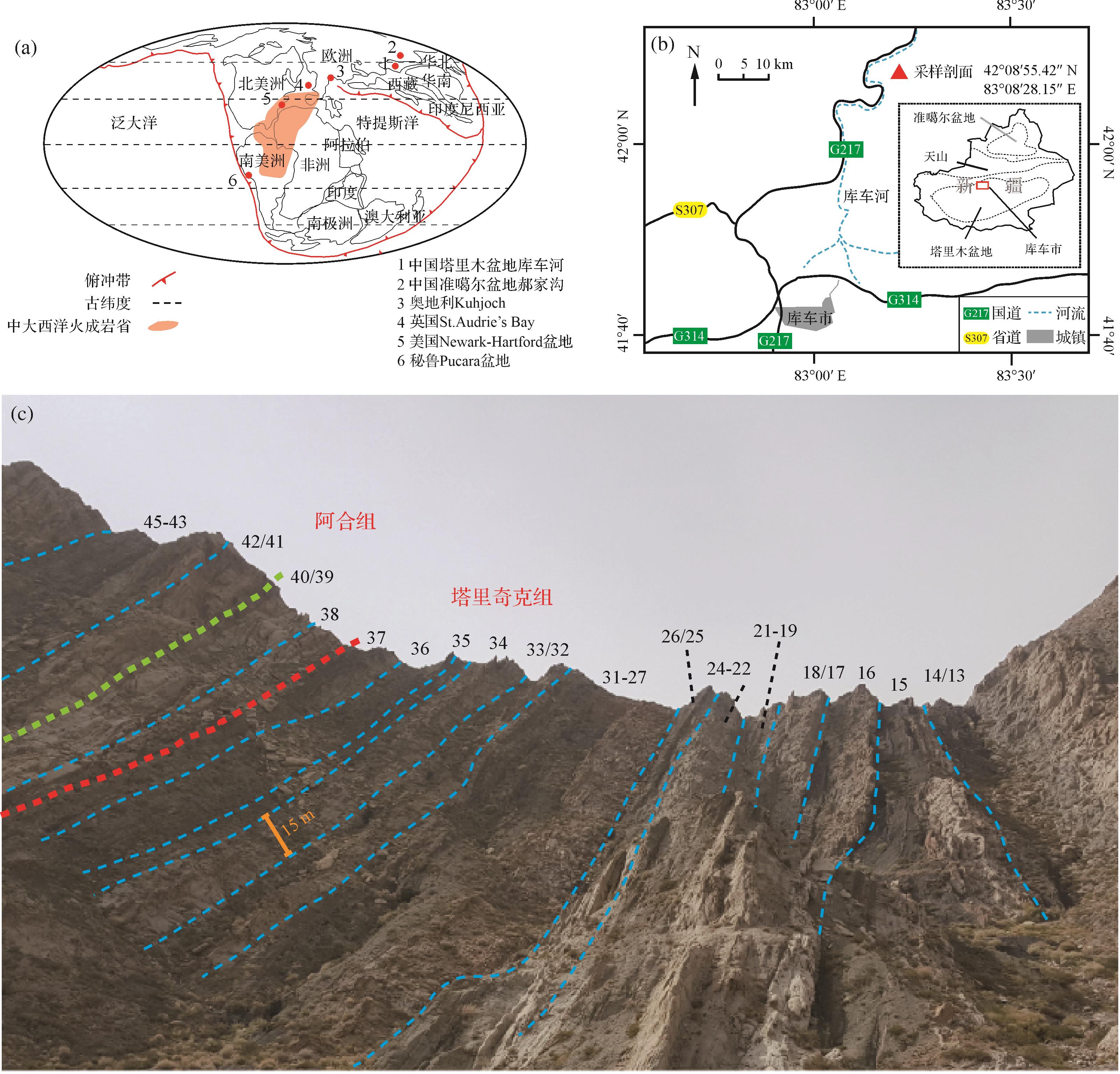
库车河剖面位于中国新疆阿克苏地区库车市北约50 km(图1),处塔里木盆地北缘,属库车坳陷构造单元。在三叠纪,古特提斯洋俯冲向北,使得盆地内中北部发育多个隆升和坳陷带,总体呈现为规模巨大的前陆盆地[60]。至侏罗纪,塔里木盆地的构造演化由新特提斯洋控制,以伸展构造、正断层为特征,形成断陷盆地[61]。在三叠纪—侏罗纪,库车河剖面位于北纬30°~40°附近(图1a)[62⁃63]。本研究目的层包括三叠系塔里奇克组以及侏罗系阿合组,13~49层(层号据王招明等[59]),厚度约270 m。地层呈单斜状产出,沉积连续无明显间断。其中13~31层属塔里奇克组下段顶部至中段,为灰色、褐灰色中厚层状含砾粗砂岩和砂岩、灰色中厚层状粉砂岩和泥质粉砂岩、黑灰色粉砂质泥岩和泥岩夹煤层组成的正韵律层。32~37层属塔里奇克组上段,为灰、褐灰色细—中砂岩和粉砂岩与深灰、灰黑色粉砂质泥岩和泥岩组成正向韵律层。38~49层属阿合组底部,为灰白色、褐灰色含砾粗砂岩、含砾中砂岩的正韵律互层,部分夹细—中砾岩[59](图1c)。

Figure 1. Paleogeographic and modern location maps, and photo of an outcrop in the Kuqa River section, Tarim Basin
综合岩性地层对比、宏体植物化石与孢粉组合,阿合组—塔里奇克组的岩性地层界线被前人厘定为该剖面的三叠系—侏罗系界线[64⁃66]。塔里奇克组发育Dictyophyllidites-Aratrisporites-Parataeniaesporites孢粉组合(DAP组合)的上亚组合;阿合组发育Disacciatrileti-Cyathidites孢粉组合(DC组合)。塔里奇克组上部和阿合组下部的孢粉组合显示了从三叠纪向侏罗纪逐步过渡的特征[59,65]。汇总的孢粉数据显示,在38层以下孢粉种属有突变发生,Alisporites auritus,Minutossaccus fusiformis,Osmuntacidites alpinus,O. elagans,O. wellmanii等种属在仍存在,至38层阿合组再未出现。同时,Asseretospora sp.,Piceites podocarpoides,Quadraeculina enigmata,Q. anellaeformis,Q. minor 在37层首现。在38层开始出现孢粉Abietineaepollenites sp., Alisporites sp., Podocarpidites sp., Piceaepollenitescomplanatiformis, P. sp., Pinuspollenites sp.[59]。以上孢粉种属数据表明,在37层与38层之间存在重要的生物延限带的界线,并与塔里奇克组和阿合组的岩性地层界线吻合,被前人厘定为三叠系—侏罗系界线[59,65],但未对ETE底界和TJB进行区分。
-
在库车河剖面,三叠系—侏罗系发育丰富的有机质碎屑,为进行陆相碳同位素地层学研究提供了宝贵的材料。本研究在库车河剖面进行了两次独立地采样工作。2010年第一批,采样从塔里奇克组13层至阿合组49层。主要采集两类样品:丝炭和煤质木化石(或其碎屑);当这两种样品不发育时,以黑色碳质页岩或者碳质泥岩作为补充。2017年第二批,采样从塔里奇克组16层至阿合组45层,采样方法与2010年相同,但第二批样品中的41~45层整体发育砂岩,所以此批样品以粉砂和细砂岩为主。两次共取得69件样品,均采集于未明显风化的岩层。选取具有代表性的丝炭和煤质木化石样品,进行光学显微镜观察和扫描电镜(SEM)观察。KQ14(04)为丝炭样品,采自塔里奇克组顶部(图2a1~a3)。KQ18(04)煤质木化石样品,采于塔里奇克组顶部(图2b1~b3)。

Figure 2. The observation of wood fossil samples under a light microscope and scanning electron microscope (SEM)
对于有机碳同位素测试的样品,将丝炭或煤化木化石约1 g装入50 mL离心管,加入容积三分之二的去离子水,用超声波清洗表面杂质,重复该步骤直至去离子水不再浑浊。之后在样品的离心管内加入3 mol/L的盐酸,55 ℃水浴中加热并震荡持续约3 h。倒掉盐酸,重复前一步实验直至加入的盐酸与样品在水浴条件下充分反应后,盐酸仍保持无色为止。之后再用去离子水在离心管内漂洗样品至pH为中性,然后在烘箱中以55 ℃放置48~72 h,至完全烘干。对于黑色碳质页岩、碳质泥岩、粉砂和细砂岩样品,用去离子水超声波清洗后烘干,粉碎至200目,称量样品4~5 g放入50 mL离心管,加入3 mol/L的盐酸在55 ℃水浴中加热并震荡,持续约3 h。后面的步骤与丝炭和煤化木化石类样品的前处理步骤相同。以上步骤除去潜在的无机碳酸根和可能破坏实验设备的硫化物。最后,用电子天平称取约50 mg样品,放入5 mm×8 mm锡杯中封装,在MAT-253稳定同位素质谱仪上进行碳同位素测试,每10个样品中加入一个平行样品和两个标样。最终获得有机碳同位素δ13C的数据结果采用PDB标准,绝对误差低于0.2‰。第一批样品在英国牛津大学地球科学系地球化学实验室完成前处理,在英国牛津大学地理系和环境系、古人类和历史艺术实验室完成测试;第二批样品在中国石油大学(北京)油气资源与探测国家重点实验室完成前处理和测试。
-
丝炭在透射光显微镜下,黑色不透明,有较高的反光度,一般长宽比大于5,呈特征性的“长条状”(图2a1)。因为丝炭在沉积埋藏前,由野火的作用下经历了无氧或缺氧条件的高温烘烤,其主体高分子成分的纤维素和木质素脱氢脱氧,同时部分支链脱落。纤维素的结构在380 ℃~400 ℃之间几乎全部遭到破坏[67],变得容易破碎,但木质素分子的维管束的支撑结构得以保留,呈现特征性长宽比的宏观以及显微结构。丝炭在扫描电镜下,可观察到原始植物细胞膜遭到破坏,但细胞壁得以保存,并且在野火缺氧烘烤的裂解作用下,其结构得以固化,在后期的压实成岩过程容易完整保存(图2a2,a3,SEM照片)。
煤质木化石在透射光显微镜下,不透明,为暗黑色,一般长宽比小于3,棱角较为清晰(图2b1)。当沉积埋藏深度小于4 m,一般为有氧条件埋藏,微生物会参与纤维素和木质素分解[68],深于4 m之后地层氧含量接近零,降解纤维素和木质素的微生物因需氧无法继续起主导作用,压实成岩作用成为主导;有机质成熟度随着地温逐渐增高,进入成煤(煤化)阶段,经历泥煤、褐煤、烟煤、无烟煤的递进过程,化学上以聚缩作用为主。煤质木化石在扫描电镜下,可见原始的微观木质结构基本被均一化压实破坏,局部偶尔保留(图2b2,b3,SEM照片)。原因是煤化成岩过程中静围岩压各方向上近似,最终形成长宽比近似的显微和宏观结构。在中生代,可以形成丝炭和煤化木化石的沉积有机物均为C3植物来源(此时C4植物还未繁盛)。根据对多块代表性样品的显微观察,显示全岩有机物中主体为保存良好的、没有强烈生物降解作用的C3植物成分。
-
2010年和2017年两次分别独立采样测试,得到库车河剖面有机碳同位素地层曲线(图3),数据见表1。根据有机碳同位素地层曲线的变化特征,从下至上划分为三段(图3)。第一段(塔里奇克组13~33层):约170 m厚,属塔里奇克组中上部。δ13Cwood主要分布范围-24.5‰至-26.5‰,总体上较稳定。第二段(塔里奇克组34~37层):约20 m厚,是塔里奇克组顶部。在此段内δ13Cwood出现了两次强烈的波动。第一次在35层底部,达2.1‰;第二次位于37层近顶部,负偏至-26.2‰后,在1 m的地层厚度内正向偏移至-22.5‰,随后急剧负偏至-26.6‰,本段内达到了4.1‰的波动幅度。第三段(阿合组38~49层):约80 m厚,为阿合组的下部。本段总体呈较轻的δ13Cwood特征,约-25.5‰至-27.4‰。并在43层底部处出现了整个剖面上的碳同位素最低值(-27.4‰);在38层顶部也记录到了一次快速负偏,幅度达1.4‰,在1 m厚范围内由-25.8‰负偏至-27.2‰。

Figure 3. Lithostratigraphy, organic carbon⁃isotope curves, and sporo⁃pollen occurrence across the Triassic⁃Jurassic boundary, Kuqa River section (data of sporo⁃pollen occurrence combined from Wang et al.[59])
样品编号 层号 厚度/m δ13Corg/(‰,PDB) 样品编号 层号 厚度/m δ13Corg/(‰,PDB) KQ10-23 46~49 81.63 -27.0 KQ2(04) 13~14 -172.10 -25.5 KQ10-22 46~49 75.32 -26.7 KQ17-y01 43~45 61.48 -25.4 KQ10-21 46~49 75.02 -26.0 KQ17-y02 43~45 57.21 -25.9 KQ10-20 46~49 71.38 -26.8 KQ17-y03 43~45 55.01 -25.7 KQ10-17 41~42 33.17 -27.4 KQ17-y04 43~45 52.71 -25.3 KQ10-16 41~42 32.17 -26.7 KQ17-y05 43~45 50.89 -26.0 KQ10-15 39~40 22.30 -26.6 KQ17-y06 43~45 43.81 -26.2 KQ10-14 39~40 21.50 -25.7 KQ17-y07 43~45 39.96 -25.4 KQ10-13 39~40 20.70 -26.0 KQ17-y08 43~45 37.14 -27.1 KQ10-12 39~40 15.90 -26.7 KQ17-y09 41~42 32.55 -26.2 KQ10-11 39~40 12.69 -26.3 KQ17-y10 41~42 31.78 -26.4 KQ10-10 39~40 12.54 -27.2 KQ17-34 41~42 29.14 -25.6 KQ10-9 39~40 12.19 -25.8 KQ17-32 39~40 22.10 -26.5 KQ17(04) 38 8.08 -25.6 KQ17-f16 38 3.00 -25.6 KQ10-7 38 6.84 -26.1 KQ17-x42(2) 38 0.30 -25.4 KQ10-6 38 4.73 -25.7 KQ17-x42 38 0.25 -24.4 KQ10-5 38 1.43 -26.1 KQ17-x41 38 0.15 -25.0 KQ10-4 38 1.03 -26.6 KQ17-x40 37 -0.35 -25.2 KQ10-3 38 0.98 -22.5 KQ17-f12 36 -8.29 -24.8 KQ10-2 38 0.78 -26.3 KQ17-f04 34 -19.92 -25.9 KQ10-1 36 -9.55 -25.1 KQ17-x39 34 -20.40 -23.6 KQ10-162 35 -18.55 -25.1 KQ17-x38 34 -22.40 -26.0 KQ10-164 34 -20.75 -24.3 KQ17-x37-1 34 -22.80 -22.5 KQ10-165 34 -22.40 -23.7 KQ17-x31 25~31 -44.54 -24.9 KQ17(04) 34 -23.28 -25.8 KQ17-x32 25~31 -45.33 -24.2 KQ10-168 32~33 -34.95 -25.8 KQ17-x29 25~31 -51.6 -25.6 KQ10-171(31-1) 25~31 -40.52 -26.0 KQ17-x25 22~24 -88.60 -26.4 KQ10-172(30-1) 25~31 -43.11 -24.7 KQ17-x18-2 19~21 -99.72 -24.0 KQ10-173(29-1) 25~31 -46.47 -26.5 KQ17-x17 17~18 -101.46 -25.7 KQ15(04) 25~31 -79.8 -25.4 KQ17-x16 17~18 -102.33 -25.5 KQ14(04) 19~21 -93.34 -25.1 KQ17-x13 17~18 -104.94 -25.8 KQ12(04) 19~21 -97.89 -25.6 KQ17-x11 17~18 -107.03 -25.2 KQ8(04) 15~16 -111.07 -26.5 KQ17-x08 15~16 -114.09 -25.3 KQ7(04) 15~16 -118.93 -25.7 KQ17-c6 13~14 -143.31 -25.7 KQ4(04) 15~16 -142.80 -25.2 2017年采样结果显示(图3),在35层内、39层下部、43层底部发生了较显著的碳同位素偏移。其中,在35层的碳同位素波动幅度达3.5‰,可与第一次采样中35层底部的δ13Cwood波动相对比;在39层下部的δ13Cwood负偏至-26.3‰,波动幅度达2.0‰,与第一次结果有较好的可重复性;43底部的δ13Cwood负偏至-27.2‰,特征同样可以与第一次结果对比,为本剖面的负偏极值。两次独立地采样的数据结果有较好的可重复性。从碳同位素曲线可以看出,以38层底为界线,之上的碳同位素整体呈现显著的负偏,平均幅度为2‰左右;38层底界恰好与地层记录中多种植物孢粉灭绝界面和首现的界面吻合(图3)。
-
TJB的GSSP以菊石Psiloceras spelae tirolicum首现面定义,即赫塘阶的底界[35],定义于奥地利Kuhjoch的海相地层剖面。该界线之下约0.5 m是孢粉组合Trachysporites⁃Heliosporites (TH)的内部两个地方性亚组带D1和D2的界线;另外菊石界线层之下3.2 m是孢粉Cerebropollenites thiergartii的首现面。TJB向下约6 m以三叠纪菊石Choristoceras marshi的末现面代表ETE底界,也恰是孢粉组合Rhaetipollis⁃Limbosporites(RL)与Rhaetipollis⁃Porcellispora (RP)的界线(图4)。

Figure 4. Biostratigraphic and carbon⁃isotope stratigraphic correlation between the Kuqa River section and global classic sections across the Triassic⁃Jurassic Boundary
英国St. Audrie’s Bay也曾是侏罗系底界GSSP候选剖面之一,是在欧洲研究较深入的海相TJB剖面。该剖面TJB位于菊石Psiloceras planorbis的首现面之下约5 m处。而牙形石灭绝带位于菊石Psilocera planorbis的首现面下7 m处,推断该层为ETE底界层位[4,77];也与蕨类植物孢子的含量“脉冲式峰值(fern spike)”的出现同步(图4)。
在美洲西部的泛大洋周缘,秘鲁的Pucara海相盆地中含有菊石Psiloceras spelae可以与TJB的GSSP剖面直接对比[30,75]。其下约5 m处为晚三叠世末菊石Choristoceras crickmayi的末现层,为ETE底界(图4)。由邻近的3个火山灰层中锆石同位素定年给出菊石Psiloceras spelae的首现面(即TJB)的绝对年龄为201.31±0.18/0.38/0.43 Ma[76](图4)。其他美洲西部的海相剖面,如美国内华达的Muller Canyon剖面的TJB由相同的菊石Psiloceras spelae首现面标定[78]。
在美洲东部的Newark裂谷盆地是TJB的经典陆相研究区,其古地理位置位于CAMP作用区内,发育了三期晚三叠世—早侏罗世的玄武岩岩浆。在该区对于湖相鱼类化石和爬行动物足迹化石研究较多,但与海相地层对比较缺乏[73⁃74]。依据陆相动物、孢粉化石以及铱元素异常数据,起初TJB被定为盆地内Orange Mountain(OM)玄武岩层之下[74]。北美东部陆相盆地的TJB与特提斯洋周缘海相剖面之间,前人进行了旋回地层学和磁性地层学对比工作。在Newark盆地与英国St. Audrie’s Bay的地磁极性地层序列对比的约束下,由天文周期旋回的短偏心率周期建立浮动天文标尺进行矫正,得到两个地区的TJB较精细的对比,表明Newark盆地的TJB应位于OM玄武岩层之上[72,79]。地层中汞浓度峰值的对比结果也显示TJB位于OM玄武岩层之上[80],这表明TJB晚于早、中期的CAMP喷发。结合美国Newark盆地和秘鲁Pucara盆地的锆石U-Pb定年结果和英国St Audrie’s Bay/East Quantoxhead旋回地层学浮动天文标尺的推算,ETE底界发生于201.58 Ma,持续约160~270 kyr至TJB[23,75⁃76];综合磁性地层学数据推断,ETE底界与TJB的间隔时长为约200~300 kyr[81]。
值得注意的是,无论是在CAMP区域的陆相盆地中(如美国Newark盆地)还是在特提斯洋西岸的海相地层中(如英国St. Audrie’s Bay地区),均在ETE底界之上紧邻TJB之下,出现了蕨类孢子含量“脉冲式”激增,指示该时段内蕨类植物迅速地、主导了陆地生态系统,但随后裸子植物快速复苏并再次占据主导(图4)。在美国Newark盆地OM玄武岩之下的ETE层处,三缝孢子含量在地层中占比从不足10%激增到超过80%;而在英国St. Audrie’s Bay地区,晚三叠世—早侏罗世孢粉产出以裸子植物孢粉为主,但是在TJB之下牙形石灭绝带之上约0.5 m处,其Selaginellales属孢子含量激增占比超过80%[77]。该现象与白垩纪末界线的蕨类爆发非常相似[73]。在GSSP的奥地利Kuhjoch剖晚三叠世末菊石Choristoceras marshi的最后出现层位(即ETE底界),主要产出少量Ovalipollis pseudoalatus,Classopollis meyeriana和Classopollis torosus孢粉组合,孢子丰度较少,属于Rhaetipollis-Limbosporites组合带。向上转变为Rhaetipollis-Porcellispora 组合带,孢子数量和种类迅速增加,蕨类和苔藓类的孢子含量达60%~80%,此组合特征持续到赫塘期早期[77]。
三叠系—侏罗系在新疆北部的准噶尔盆地南缘发育非常连续,郝家沟剖面是典型的陆相TJB剖面[82⁃83]。郝家沟剖面植物组合和孢粉组合在郝家沟组和八道湾组之间变化明显,在郝家沟组内Danaeopsis植物及Aratrisporites和Chordasporites等孢粉指示晚三叠世,八道湾组发育Coniopteris gaojiatianensis等指示早侏罗世[84],因此TJB位于郝家沟组顶部与八道湾组底部之间[82,84]。另外,在郝家沟组上部43层开始也有类似的蕨类孢子含量激增现象[85],可与奥地利Kuhjoch、英国St. Audrie’s Bay和美国Newark盆地的三个典型剖面的ETE事件层对比(图4)。
在库车河剖面尚未有孢粉丰度数据,但塔里奇克组37层有多种蕨类孢子和裸子植物花粉的首现,如Asseretospora sp., Piceites podocarpoides,Quadraecu lina anellaeformis,Q.enigmata,Q.minor,等[59],暗示较大规模的、陆相植被生态系统的波动;推测可对比于郝家沟剖面43~45层,并可对比于奥地利Kuhjoch、英国St. Audrie’s Bay和美国Newark盆地的剖面(图4)。综合库车河剖面生物地层及孢粉统计资料,图4中汇总了与经典剖面的ETE底界和TJB层位的对比。综上,综合的地层学研究成果为库车河剖面进行跨海、陆精细的地层厘定和对比提供了良好的约束。
-
在4.1部分讨论的地层框架约束基础上,对库车河剖面与GSSP剖面及其他经典剖面进行了碳同位素地层学精细对比。在奥地利Kuhjoch的GSSP剖面,全岩有机质碳同位素(δ13Corg)曲线显示I-CIE和M-CIE的显著特征[25]。在剖面约0 m处出现了一次短暂的-7‰CIE,即为I-CIE,与晚三叠世末菊石Choristoceras marshi末现面一致,即ETE底界。在δ13Corg迅速恢复之后,在剖面约6 m处呈现幅度达-3‰的负偏,其在M-ICE之下与侏罗纪菊石Psiloceras spelae tirolicum首现面一致,即TJB(图4)。可以指示陆源高等植物碳源的长链正构烷烃的碳同位素(δ13Calk)地层曲线与全岩有机碳结果相似,I-CIE幅度为-6‰[26]。在英国St.Audrie’s Bay剖面,I-CIE位于Cotham组与Langport组交界,牙形石带顶部,呈现约4‰的短暂负偏;I-CIE上方约5 m处为M-CIE,幅度约为2‰,保持相对轻的碳同位素值,一直持续至辛涅缪尔期,菊石Psiloceras planorbis的首现面在此附近[4]。美国的Newark盆地的湖相地层中记录了单体正构烷烃、陆源木质碎屑以及全岩有机质碳组分[27],在该剖面三叠系顶部的δ13Calk负偏幅度达4‰;随后δ13Calk正偏幅度达5‰。在郝家沟剖面,δ13Corg地层曲线在42层、43层与45层之间呈现约1.5‰~2‰的CIE;结合孢子激增层位、晕苯和汞元素峰值出现的层位,推断I-CIE位于43层底部附近;49~51层底部附近出现约2‰的M-CIE,此后地层向上为相对长期的、稳定的正向偏移趋势[69,85⁃86]。
在库车河剖面,塔里奇克组38层底部附近(约0 m处)、39层底部附近(约10 m处)和43层底部(约30 m处),分别出现了三次碳同位素负偏,幅度约1.7‰~2.5‰(图3)。碳同位素地层曲线负偏特征显著,推断I-CIE位于38层底部附近。另外,宏观的沉积环境在38层附近有较大转变,从沼泽和河流为主的沉积环境变为以较大规模的河流相沉积为主,以煤线或薄煤层为特征的沼泽相沉积在38层之上不再出现(图3)。38层也开始记录到新孢子种属的出现,暗示可与地层中蕨类孢子激增事件对比;结合其他生物年代地层约束(图4),本研究认为ETE底界位于塔里奇克组与阿合组之间的岩性地层界线,即38层底部附近。从43层底部附近出现的δ13Cwood约2.5‰负偏,以及此层位之上开始的相对长期的、稳定的正向偏移趋势,可以推定此为M-CIE起始层位;而全球海陆地层对比的结果明都确显示,以海相菊石首现面定义的TJB位于M-CIE之下的次一级碳同位素负偏层位(图4),综合孢粉记录Piceaepollenites exlioides,Asseretospora spp.和Osmuntacidites sp.等种属在39~43层的首现(图3),暗示生态系统的逐步恢复,推论本剖面的TJB位于41层底部附近。
-
在美国内华达洲的海相TJB,在准噶尔盆地和四川盆地的陆相TJB附近,均观测到异常高的、幔源的汞元素丰度[80,86⁃87],表明CAMP虽然直接作用区集中在现今非洲西北部、北美洲东部和南美洲北部,其影响的规模可达全球。在南美洲秘鲁的Pucara盆地,三叠系—侏罗系发现多层火山灰层,取得了锆石年代学结果[76]。在摩洛哥Argana盆地最早的CAMP玄武岩年龄老于Newark盆地的[88],由于Newark盆地的玄武岩层是直接覆盖于该盆地的ETE事件层之上,可认为与灭绝事件近同期,因此表明ETE的发生在CAMP的大规模喷发之后[81,88]。
晚三叠世末全球多处海相和陆相地层中,同期都记录了碳同位素地层曲线正、负偏的显著波动,根据基于C3植物的碳同位素地层学理论,推论该时期大气中CO2的δ13C亦发生显著波动,指示全球性的、地表圈层的碳储库的波动。CAMP大火成岩省的岩浆作用,导致大量幔源碳释放到大气中,迅速地使大气CO2浓度增加2~4倍[8,10⁃11,89⁃90]。但CAMP爆发释放的幔源δ13C值为-5‰左右,与大气CO2的δ13C长期稳定值-5‰~-7‰相近(或略偏重),无法引起负向约2‰的偏移;2‰的负偏仅可能源于外来轻碳同位素源注入大气,或者海、陆初级生产者遭遇毁灭性打击的结果——初级生产者优先固定轻碳同位素的生物泵作用全球规模地停摆;或者此两种原因混合。
轻碳同位素主要来源有潜在的三种情况之一,或其不同比例组合:第一,因板块构造裂解,造成的大陆架上失稳的生物成因CH4气水合物释放(特征δ13C值约-60‰)到大气中,其温室效应约是CO2的25倍,约30年内CH4全部转化成CO2[4,54];第二,全球性的森林火灾,植被(特征δ13C值约-26‰)燃烧释放到大气中的CO2[24];第三,因全球升温,导致的南北极高纬度地区的或高海拔地区的永久冻土带中封存的生物成因甲烷气水合物(特征δ13C值约-60‰)释放到大气中[91⁃92]。以上一种或多种轻碳源注入大气圈后,使得在海水的碳酸盐岩中和C3植物中的碳同位素值呈现约2‰负偏的记录,呈现全球性的地球表层系统碳库的波动。未来对CAMP的玄武岩中挥发性物质的定量研究,以及其如何影响全球生态系统并导致海陆生态系统的崩溃,可能是解开三叠纪末期一系列关键地质事件的钥匙。该事件的定量研究成果,对当代全球碳库波动模型的建立和验证参数的优化,具有实证数据的借鉴意义。
-
(1) 在新疆塔里木盆地北缘的库车河剖面,两次独立地有机碳同位素地层学样品采集和测试,其结果具有较好的可重复性。
(2) 在库车河剖面完成了跨海、陆的TJB碳同位素地层学对比。推断与全球性的ETE事件等时底界,位于38层底部附近;与GSSP等时的TJB地层界线,位于41层底部附近;ETE底界与TJB的间隔时长为约200~300 kyr。
(3) 在ETE底界和TJB附近,陆相和海相的碳同位素地层曲线都有2‰~7‰幅度不等的负偏。I-CIE(即ETE底界)与地层中孢粉含量“脉冲式”爆发事件的起始层位同期,表明蕨类植物响应于ETE,其短暂地占据陆地生态系统优势生态位,暗示生态系统快速剧烈波动。M-CIE的起始临近层位对应海相菊石首现定义的TJB,此时的首现物种暗示全球生态系统复苏的开始。
(4) 全球多处海相和陆相地层中,同期都记录了碳同位素地层曲线正、负偏的显著波动,推论该地质时期大气中可能有轻碳源注入或者初级生产者遭遇毁灭性打击,或两者共同作用,导致了全球性的、地表圈层的碳储库的波动。

 DownLoad:
DownLoad:


